
Abstract
SUMMARY: In clinical MR spectroscopy at higher field strengths, lactate may show reduced or absent signal intensity at an echo time of 144 ms. Although this false-negative result may be predicted from theory, experimental verification and clinical impact have not been fully established. Using scanners from 3 major vendors, spectra from phantoms and patients demonstrate the lactate signal loss and potential error in interpretation. Strategies are discussed to overcome, or at least alleviate, this problem.
Lactate is a metabolite that plays a pivotal role in many brain pathologies such as tumors, stroke, cerebral ischemia, hypoxia, and several mitochondrial disorders.1 The lactate concentration in healthy brain under normal conditions is approximately 1 mmol/L. However, when oxygen availability is low because of metabolic stress, glucose molecules are no longer oxidized completely and pyruvate is produced. The pyruvate is converted to lactate, which can rise to concentrations >10 mmol/L. Lactate is, therefore, an important marker of anaerobic glycolysis taking place in the previously mentioned brain disorders.
The lactate molecule has 2 weakly coupled resonances in 1H-MR spectroscopy: a doublet (split by coupling to the methine [CH] proton) at 1.33 ppm, arising from 3 magnetically equivalent methyl (CH3) protons, and a quartet (split by coupling to the protons of the methyl group) at 4.11 ppm, arising from the methine proton, which is usually not visible in vivo. The scalar coupling gives rise to a phase evolution of the methyl doublet, which depends on the echo time (TE). For TE = 144 ms, the resonance shows a phase of 180° leading to a negative in-phase doublet, whereas TE = 288 ms gives rise to a positive in-phase doublet. Since only in-phase resonances can be quantified, echo times of 144 ms and 288 ms are preferable for lactate detection and assignment. The coupling evolution can also be exploited for spectral editing techniques, increasing the sensitivity of lactate detection.2
Localization techniques such as the double spin-echo method called point-resolved spectroscopy (PRESS), which is the standard localization technique used on clinical MR imaging systems, cause chemical shift displacement artifacts. This can give rise to signal misregistration for almost all metabolites because only the signal from one specific frequency, usually the N-acetylaspartate (NAA) frequency, originates from the selected volume of interest. Signal from protons with different chemical shifts (eg, from other metabolites) stem from spatially shifted volumes. Furthermore, for weakly coupled resonances, there is an additional signal cancellation due to anomalous J-modulation.3, 4 This additional artifact and its effect on the interpretation of lactate levels in clinical spectra are discussed in this article. Since the chemical shift displacement roughly scales with the square of the field strength (in part because of reduced radio-frequency [RF] pulse bandwidths, as well as increased chemical shift frequency separations), a severe underestimation of lactate occurs at 3T, when PRESS localization is used with TE = 144 ms. A detailed explanation of the origin of anomalous J-modulation can be found in the Appendix.
This article shows in vitro and in vivo examples of signal cancellation for several MR imaging systems. A strategy is presented to quantify the lactate signal loss and thus validate the underlying theory. Finally, suggestions for parameter choices on clinical systems are given to avoid or at least diminish the problem of lactate underestimation at 3T.
Description of the Technique and Results
Patient Measurements
Two patients with high-grade gliomas underwent MR spectroscopy, both with echo times of 144 and 288 ms, performed on Philips Intera whole-body systems (Philips Medical Systems, Best, The Netherlands) by using a send-receive head coil. One patient was measured at a field strength of 1.5T, whereas the other was measured at 3T.
Another patient with mitochondrial myopathy, encephalopathy, lactic acidosis, and stroke (MELAS) was examined on 1.5T and 3T GE Signa systems (GE Healthcare, Milwaukee, Wis) from a second vendor. Two-dimensional multivoxel spectra were acquired at both field strengths consecutively the same day, repeated with both TE = 144 ms and TE = 288 ms. Standard PRESS localization was used with the region of interest centered over an acute occipital lesion. On this system, very selective saturation (VSS) bands6 are placed around the volume of interest by default.
Some of the data presented here were acquired on a clinical basis, but all patients gave their written informed consent before participating in research experiments.
In the high-grade tumor in Fig 1, the spectra acquired at 1.5T both show a prominent lactate doublet, inverted for TE = 144 ms and in-phase with NAA for TE = 288 ms. The lactate peak, clearly visible as an inverted doublet at TE = 144 ms, is of greater area and amplitude at TE = 144 ms than at TE = 288 ms. This signal change reflects the known T2-related signal attenuation with increasing echo times.

Single-voxel spectra acquired at 1.5T from the brain of a patient with a high-grade glioma by using PRESS localization. B, TE = 144 ms. C, TE = 288 ms. Cho indicates choline; Lac, lactate; NAA, N-acetylaspartate.
Comparable experiments were carried out on a 3T system, scanning another patient with a glioma (grade III). Although for TE = 288 ms a large lactate peak is observable at 1.33 ppm, the lactate resonance has completely vanished for TE = 144 ms due to anomalous J-modulation (Fig 2).
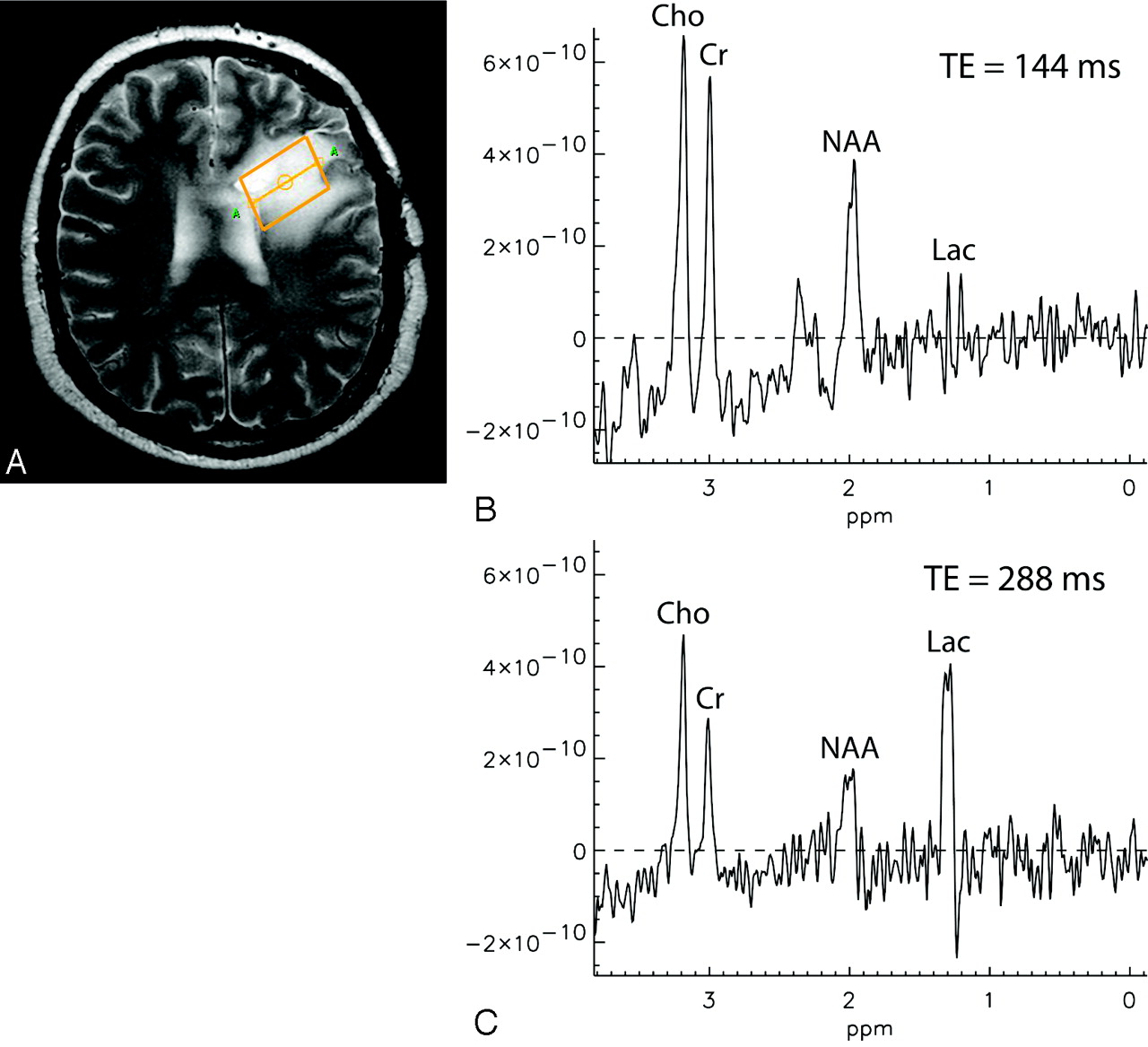
Single-voxel spectra acquired at 3T from the brain of a patient with a grade III glioma by using PRESS localization. B, TE = 144 ms. C, TE = 288 ms. Cr indicates creatine.
The third patient example (Fig 3), carried out on different systems, shows lactate peaks of similar relative amplitude with TE = 144 ms and TE = 288 ms at 1.5T. The lactate signal loss, which is expected with TE = 288 ms compared with TE = 144 ms because of T2 relaxation, is approximately compensated by the signal loss due to anomalous J-modulation with TE = 144 ms. At 3T, however, anomalous J-modulation gives rise to a complete disappearance of the lactate peak.

Multivoxel spectra acquired 1 hour apart at 1.5T and 3T from the same region in the brain of a patient with MELAS, by using standard PRESS localization with TE = 144 ms and TE = 288 ms. An inverted lactate doublet is clearly visible at 1.5T, but not at 3T (arrows). Upright lactate peaks at TE = 288 are seen equally well at both field strengths (arrows).
Phantom Measurements
A standard brain metabolite phantom containing 5 mmol/L of lactate was measured on 3 3T MR imaging scanners from 3 different vendors (GE Healthcare, Philips Medical Systems, and Siemens Medical Solutions). Single-voxel MR spectra (volume of interest [VOI] size = 2 × 2 × 2 cm3, PRESS localization) were acquired from the same volume once with TE = 144 ms and once with TE = 288 ms. The lactate peak seen at 1.33 ppm for TE = 288 ms would be expected to be smaller than the inverted doublet for TE = 144 ms as a result of T2 relaxation. However, at 3T, spectra from all 3 MR systems show significantly larger lactate resonances at TE = 288 ms than at TE = 144 ms. For one system, the lactate peak has even disappeared completely. This disappearance demonstrates that a significant amount of the lactate signal is lost (not visible) at TE = 144 ms (Fig 4).

Proton spectra acquired from a standard brain metabolite phantom containing 5 mmol/L of lactate. The measurements were performed on 3 3T MR imaging scanners from 3 different vendors (Philips Medical Systems, GE Healthcare, and Siemens Medical Solutions). Single-voxel MR spectra (VOI size = 2 × 2 × 2 cm3, PRESS localization) were acquired from the same volume once with TE = 144 ms and once with TE = 288 ms. Radio-frequency pulse bandwidths for the selective refocusing pulses vary between vendors in the range of 874–2300 Hz.
To quantify the signal loss due to anomalous J-modulation on the basis of the phantom spectra acquired for TE = 144 ms and TE = 288 ms, one would have to determine the T2 relaxation constant of lactate first. Since the relaxation constant of lactate for the used brain metabolite phantom was unknown and difficult to determine with high precision, a different approach was chosen: MR spectroscopic imaging (MRSI) offers the possibility of dispensing with spatially selective refocusing pulses because the MRSI slice can be selected only by the excitation pulse, and the spatial encoding within the slice is achieved with phase-encoding only. However, in a standard MRSI measurement protocol without PRESS localization, both the excitation pulse and the refocusing pulse, which is needed for the echo formation, are slice-selective on some scanners, leading again to some signal loss. To estimate the amount of signal loss due to anomalous J-modulation, we acquired MRSI datasets once with (“standard” MRSI sequence) and once without the slice-selection gradient during the refocusing pulse. Theory predicts that the overall signal loss at TE = 144 ms is the same for a standard MRSI sequence as for a single-voxel protocol using PRESS (Appendix). It can also be calculated that no signal loss should occur for lactate for a standard MRSI measurement with TE = 288 ms. Therefore, to validate the theoretic predictions experimentally, MRSI datasets were acquired for these 2 echo times. Postprocessing of the spectra included exponential filtering of the time domain signal cosine filtering in k-space, and B0 correction.
Figure 5 shows the results of the MRSI measurements without PRESS. For echo times of 144 ms and 288 ms, one representative MRSI voxel is shown (acquired once with [Figs 5A, -C] and once without a refocusing pulse gradient [Figs 5B, -D], respectively).

Spectra from an MRSI imaging dataset acquired at 3T from a phantom containing 10 mmol/L of NAA and 20 mmol/L of lactate without PRESS localization. A, TE = 144 ms, with refocusing pulse gradient. B, TE = 144 ms, without refocusing pulse gradient. C, TE = 288 ms, with refocusing pulse gradient. D, TE = 288 ms, without refocusing pulse gradient.
Integrating the modulus spectra and comparing the results with and without anomalous J-modulation yielded a signal loss of 72.2% for TE = 144 ms, whereas the signal intensity was approximately the same for TE = 288 ms. Theoretic calculations taking into account the chemical shift difference of the coupled nuclei and the bandwidth of the used refocusing pulses yield a relative signal loss of 81.2% for TE = 144 ms, whereas for TE = 288 ms, no signal loss due to anomalous J-modulation is predicted (Appendix).
Thus the measured signal loss is consistent with the theoretically expected value. In contrast to MRSI without PRESS, theory also predicts a small signal loss of 6.8% at TE = 288 ms when using PRESS localization for single-voxel experiments. However, this loss only needs to be taken into account when high-precision quantification is desired.
Discussion
Detection of lactate using MR spectroscopy plays an important clinical role in the assessment of a number of brain abnormalities, including tumor, stroke, and mitochondrial disorders. The presence of lactate in the context of a tumor can be considered diagnostic for glioblastoma multiforme. Lactate is also elevated as a consequence of mitochondrial abnormalities in neurodegenerative disorders such as Huntington disease. Since for very short echo times, there are often residual fat peaks visible in the spectral region of the lactate doublet, one has to resort to echo times near multiples of 1/J (J being the coupling constant) for proper lactate detection and quantification. As an inverted doublet can be discriminated more easily against other resonances like lipids, PRESS with TE = 144 ms is often considered the most appropriate method for unambiguous lactate detection at 1.5T. Although the effect of anomalous J-modulation and the potential signal loss for lactate at TE = 144 ms have been discussed in the literature,3, 4 little attention has been given to this phenomenon in clinical routine so far. Neglecting this effect can potentially lead to a severe misinterpretation in clinical diagnosis as shown in our examples. The extent of the signal loss due to anomalous J-modulation can vary considerably depending on the field strength, the used coil, and the sequence parameters. A practical recommendation for clinical MR spectroscopy at 3T is to perform a phantom study at TE = 144 ms and TE = 288 ms; if the lactate signal intensity at TE = 144 ms is less than that seen at TE = 288 ms, it is recommended not to use TE = 144 ms, but rather only TE = 288 ms in clinical examinations, although in general, the sensitivity decreases with longer echo times because of T2 relaxation. However, the quantitative influence of anomalous J-modulation can be different in specific in vivo examples, in which inhomogeneous metabolite distributions can either aggravate or attenuate the effect compared with in vitro experiments.
Strategies to prevent or alleviate the signal loss due to anomalous J-modulation are discussed elsewhere in detail3–5 but usually require changes in the scanner software. An approach implemented on several clinical scanners is to saturate the region of spin-selective refocusing with outer volume suppression pulses before excitation. For example, on some scanners, quadratic-phase suppression pulses have been implemented for this purpose,6 but they suppress the effect only partially, because they are not specifically tuned for lactate detection as can be seen both from in vivo and phantom examples in this article. However, at higher field strengths than 3T, which have now become available for human studies as well, the chemical shift displacement will be so pronounced that this work-around will be less effective.
Another approach for high-field systems is using pulses with much larger bandwidths, such as adiabatic pulses, to alleviate the chemical shift displacement. However, since ordinary adiabatic pulses are usually not spatially selective, they cannot be used in PRESS sequences. Localization by adiabatic selective refocusing (LASER) applies pairs of large-bandwidth adiabatic full passage pulses for volume selection and echo formation at the same time and will probably be the method of choice for localization in high-field spectroscopy applications in the future.7 A further popular localization technique is STimulated Echo Acquisition Mode (STEAM), which uses 3 90° pulses giving rise to a stimulated echo. Since 90° pulses have much larger bandwidths than 180° pulses, the chemical shift displacement is far less severe. However, for STEAM, the signal intensity of coupled resonances not only varies with TE but also shows a strong modulation governed by the mixing time between the second and third pulse.1 Furthermore, MRSI offers the possibility to dispense with spatially selective refocusing pulses and, therefore, prevent the signal cancellation. However, MRSI protocols without PRESS localization are usually not implemented on purely clinical scanners, and very good outer-volume suppression is required for this approach to prevent the spectra from being impaired by subcutaneous fat signal.
Appendix
The signal cancellation by anomalous J-modulation for coupled resonances arises from the chemical shift displacement artifact. The relative voxel displacement for 2 protons with chemical shifts δ1 and δ2 equals the ratio of the chemical shift difference (ΔωCS = δ1 –δ2) and the bandwidth of the RF pulse (ΔωRF) used for volume selection. For the signal intensity of the methyl (CH3) resonance of lactate at 1.33 ppm, the signal loss can be understood as follows: Due to this chemical shift displacement, the volume selected by one single refocusing pulse decomposes into a region in which both the CH3 and the methine protons of the lactate molecule are affected by the pulse (“nonselective” pulse) and a region in which only the methyl spin is inverted (“selective” pulse), and therefore the coupling evolution is refocused. This leads to a superposition of signal with different phases giving rise to signal cancellation in the spectrum. The volume selected by a PRESS sequence, which uses 2 refocusing pulses, consists of 4 partial volumes with different phase evolutions, depending on whether none, one, or both of the 2 refocusing pulses are spin-selective (Fig 6). In a PRESS sequence, the signal loss is also determined by the sequence timing because the spins are inverted twice by the 2 spatially selective refocusing pulses.8 By adding the signal terms for the 4 partial volumes, one can calculate the resulting signal loss. This effect, termed “anomalous J-modulation,”4 is particularly pronounced for TE = 144 ms, when the 2 superimposed signals from the partial volumes V1 and V4 have a phase difference of 180°, and therefore the cancellation is most effective, whereas for TE = 288 ms, almost no cancellation occurs (Fig 6).

Partial volumes and their coupling evolution for a single-voxel PRESS experiment: The 90° excitation pulse is applied with a gradient in the z direction, whereas the 2 refocusing pulses are applied with gradients in the x and y direction, respectively. The size of the partial volumes is determined by the chemical shift displacement. The signal phase of the magnetization is determined by TE, the scalar coupling constant (J) of lactate, and the time interval (t1) between the excitation pulse and the first refocusing pulse.
According to theory, the relative signal loss for lactate due to anomalous J-modulation at TE = 144 ms amounts to 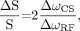 where ΔωCS is the chemical shift difference of the coupled nuclei and ΔωRF is the bandwidth of the refocusing pulse. At 3T the chemical shift difference between the methyl and the methine proton is 355 Hz and the refocusing pulse used in this example had a bandwidth of 874 Hz. This predicts a relative signal loss of 81.2% due to anomalous J-modulation for TE = 144 ms. The theoretic signal loss for single-voxel PRESS measurements at TE = 288 ms depends on the position of the 2 refocusing pulses within the sequence. Usually the PRESS sequence is rendered as asymmetric as possible, with the first refocusing pulse being irradiated as soon as possible (at time t1) after the excitation pulse. For the standard PRESS sequence implemented on a 3T Philips Intera scanner, theory predicts the following signal loss due to anomalous J-modulation at TE = 288 ms:
where ΔωCS is the chemical shift difference of the coupled nuclei and ΔωRF is the bandwidth of the refocusing pulse. At 3T the chemical shift difference between the methyl and the methine proton is 355 Hz and the refocusing pulse used in this example had a bandwidth of 874 Hz. This predicts a relative signal loss of 81.2% due to anomalous J-modulation for TE = 144 ms. The theoretic signal loss for single-voxel PRESS measurements at TE = 288 ms depends on the position of the 2 refocusing pulses within the sequence. Usually the PRESS sequence is rendered as asymmetric as possible, with the first refocusing pulse being irradiated as soon as possible (at time t1) after the excitation pulse. For the standard PRESS sequence implemented on a 3T Philips Intera scanner, theory predicts the following signal loss due to anomalous J-modulation at TE = 288 ms: 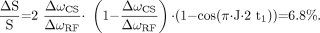 For an MRSI sequence without PRESS localization but with a slice-selective refocusing pulse for the echo formation, there are only 2 interfering partial volumes (corresponding to V1 and V4 in Fig 6). For TE = 144 ms, this leads to the same relative signal loss as for a single voxel PRESS sequence, whereas no signal loss at all occurs for TE = 288 ms.
For an MRSI sequence without PRESS localization but with a slice-selective refocusing pulse for the echo formation, there are only 2 interfering partial volumes (corresponding to V1 and V4 in Fig 6). For TE = 144 ms, this leads to the same relative signal loss as for a single voxel PRESS sequence, whereas no signal loss at all occurs for TE = 288 ms.
To understand the basic behavior of the lactate resonances in a PRESS sequence, one can approximate the used pulses as ideal hard pulses. However, pulse imperfections give rise to a far more complicated evolution behavior, which was investigated analytically, simulated numerically, and discussed in detail in several publications.9–13 Since the chemical shift is proportional to B0 and, because of B1 limitations, the pulse bandwidths are approximately inversely proportional to B0, the chemical shift displacement and therewith the lactate signal loss due to anomalous J-modulation at TE = 144 ms roughly scale with the square of the field strength.
Acknowledgments
We thank Drs. Andy Alexander and Karl Vigen, University of Wisconsin, Madison, and Dr. D-J Wang, CHOP, Philadelphia, for helping with the acquisition of some of the spectra on the scanners of different vendors.
Footnotes
This work was supported by grant SEP TH 7/02–2, ETH Zurich, Zurich, Switzerland.
Presented at the 43rd annual meeting of the American Society of Neuroradiology, Toronto, Ont, Canada, May 2005.
- Received July 25, 2005.
- Accepted after revision November 16, 2005.
- Copyright © American Society of Neuroradiology
In this issue









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Spatiospectral image processing workflow considerations for advanced MR spectroscopy of the brain
- Does Gadolinium Deposition Lead to Metabolite Alteration in the Dentate Nucleus? An MRS Study in Patients with MS
- Lower Lactate Levels and Lower Intracellular pH in Patients with IDH-Mutant versus Wild-Type Gliomas
- Expression Changes in Lactate and Glucose Metabolism and Associated Transporters in Basal Ganglia following Hypoxic-Ischemic Reperfusion Injury in Piglets
- Utility of Proton MR Spectroscopy for Differentiating Typical and Atypical Primary Central Nervous System Lymphomas from Tumefactive Demyelinating Lesions
- Focal MR spectroscopy of hippocampal CA-1 lesions in transient global amnesia