
Abstract
BACKGROUND AND PURPOSE: The cerebral and cerebellar networks involved in bimanual object recognition were assessed by blood oxygen level–dependent functional MR imaging by using multivariate model-free analysis, because conventional univariate model-based analysis, such as the general linear model (GLM), does not allow investigation of resting, background, and transiently task-related brain activities.
MATERIALS AND METHODS: Data from 14 healthy right-handed volunteers, scanned while successively performing bilateral finger movements and a bimanual tactile-tactile matching discrimination task were analyzed by using tensor-independent component analysis (TICA), which computes statistically independent spatiotemporal processes (P > .7) thought to reflect specific and distinct anatomofunctional neural networks. These results were compared with the network obtained in a previous study by using the same paradigm based on GLM to evaluate the advantages of TICA.
RESULTS: TICA characterized and distinguished the following: 1) resting-state networks such as the default-mode networks, 2) networks transiently synchronized with the beginning and end of the task, such as temporo-pericentral and temporo-pericentral-occipital networks, and 3) task-related networks such as cerebello-fronto-parietal, cerebello-prefrontocingulo-insular, and cerebello-parietal networks.
CONCLUSION: Bimanual tactile-tactile matching discrimination specifically recruits a complex neural network, which can be dissociated into 3 distinct but cooperative neural subnetworks related to sensorimotor function, salience detection, executive control, and, possibly, sensory expectation. This tripartite network involved in bimanual object recognition could not be demonstrated by GLM. Moreover, TICA allowed monitoring of the temporal succession of the networks recruited during the resting phase, audition of the “go” and “stop” signals, and the tactile discrimination task.
In a previous study,1 we found that bimanual tactile-tactile matching discrimination of small objects recruited multiple sensorimotor and associative cerebral and neocerebellar areas. However, this neural network may represent a combination of cooperative but functionally distinct subnetworks and could also underestimate the total number and identity of the networks actually recruited by this type of sensorimotor task because functional data were processed by the very restrictive general linear model (GLM). This univariate approach only identifies task-related effects whose time-course is well correlated with the expected hemodynamic changes and merges them on a final unique statistical parametric map. Therefore, independent concomitantly activated networks cannot be clearly dissociated, and transiently task-related networks cannot be detected. To circumvent these major limitations, one must use multivariate model-free data-analysis techniques, such as probabilistic independent component analysis,2 to determine statistically relevant complex time-modulated responses in functional MR imaging (fMRI) data. This method can also provide useful access to networks activated during the rest phase, some of which could specifically participate in the brain resting state,3–7 whereas others could participate in preparatory sensorimotor or cognitive processes linked to the following activity phase.
We, therefore, reassessed the neural networks involved in the bimanual tactile-tactile discrimination task. More precisely, we determined the following: 1) whether the network previously described1 must be divided into several different subnetworks, 2) the temporal order of recruitment of these possible circuits, 3) whether other relevant networks were not previously identified by the GLM method, and, incidentally, 4) whether low-volume sampling (ie, short data-acquisition sequences) can provide significant results without aliasing. This complementary study, therefore, applied a probabilistic tensorial extension of independent component analysis for multisubject sessions to blood oxygen level–dependent fMRI (BOLD fMRI) data.2
Materials and Methods
Subjects
This analysis was performed on 14 healthy right-handed subjects (range, 19–40 years of age; mean, 26.5 years of age), all of whom gave their informed consent before the study. These subjects had no history of cardiovascular, neurologic, or psychiatric disease.
Task Procedure
As previously described,1 subjects lay supine with closed eyes and both hands extended in an MR imaging machine (Signa Horizon, GE Healthcare, Milwaukee, Wis). Cerebral and cerebellar activation was studied during the following bimanual tasks: 1) bilateral finger movements, and 2) a tactile-tactile matching task, with no visual input. The fMRI session consisted of 9 alternating 15-second phases: rest (nonmovement rest condition)/motor task. For each task, the total procedure lasted 2.25 minutes. Before scanning, each hand was placed in a separate bag, and the 2 bags contained identical sets of chess pieces (10 × 5 × 5 mm). During the motor phase, subjects were asked to grasp a chess piece in each hand and to drop the pieces before repeating the procedure. During the sensorimotor task, subjects were asked to grasp a chess piece in each hand, compare these chess pieces by palpation to determine whether they were identical or different, and drop the pieces before repeating the procedure. As in the motor task, palpation involved the thumb, index finger, and middle finger.
fMRI Data Acquisition
fMRI was conducted on a whole-body 3T clinical imager (Signa Horizon) by using an 8-channel head coil. In each scanning sequence, 32 contiguous axial T2*-weighted gradient-echo echo-planar images (TE/TR, 40/2500 ms; FOV, 30 × 30 mm; matrix, 128 × 128; thickness, 4 mm; intersection spacing, 0 mm; NEX, 1) were obtained to encompass the entire brain and cerebellum. Fifty-eight volumes were acquired including 4 “dummy” volumes obtained at the start of the session. Off-line, T2*-weighted images were coregistered and overlaid on their corresponding anatomic inversion-recovery–weighted gradient-echo images (TE/TI/TR/flip angle, 3/450/7.3 ms/20°; FOV, 25.6 × 25.6 mm; matrix, 256 × 256; thickness, 1 mm; intersection spacing, 0 mm).
Statistical Analysis
Functional image analysis used FSL, Version 4 (FMRIB Software Library, http://www.fmrib.ox.ac.uk/fsl/). Data were successively format-converted, motion-corrected (MCFLIRT, www.fmrib.ox.ac.uk/fsl/mcflirt/index.html), spatially smoothed with a 5-mm full width at half maximum of Gaussian kernel, intensity-normalized, high-pass temporal filtered, and coregistered by using the template brain of the Montreal Neurologic Institute (MNI) proportional spatial scaling. Multisubject tensor-independent component analysis was then applied to these preprocessed data (Multivariate Exploratory Linear Optimized Decomposition into Independent Components). As previously explained,2,7 these data from all 14 volunteers were decomposed into independent maximally non-Gaussian components (IC), whose number was determined by principal component analysis. Spatial maps associated with each IC were transformed to z-scores. For each resulting Z-map, a combined Gaussian/γ model of the probability attenuation of background noise was then applied to the distribution of voxel intensities to determine which of these voxels was significantly modulated by the associated time course, with a posterior probability threshold (of activation) of P > .7. Clusters were considered significant whenever they included more than 4 voxels. Both spatial maps and time courses were constrained to be the same for all subjects. These component or activation maps were overlaid on the mean T2 image in the stereotactic MNI152 standard space. Moreover, comparison of the time series with the paradigm design consisting of alternating rest/activity phases allowed determination of the following: 1) sequential recruitment of the associated network across the whole paradigm, 2) relative synchronization of the recruited networks, and 3) correlation of networks with the task. This statistical analysis was performed separately for the motor and sensorimotor tasks.
Results
Tensor-Independent Component Analysis
BOLD fMRI data during the motor and tactile discrimination tasks were analyzed separately by using tensor-independent component analysis (TICA). According to the preliminary eigen spectrum analysis of the principal component analysis, 15 and 18 independent components were retained for the motor and discrimination tasks, respectively. However, components for which the time course was not clearly task-related, with insufficient explained variance (>5%), or which corresponded to known artifacts such as movements or noise8 were identified by visual inspection and rejected by 1 rater.
Nondiscrimination Motor Task
Because this task was performed only to obtain the components related to the motor networks, the other (nonmotor) components were not studied further.
Component 1: Motor Network.
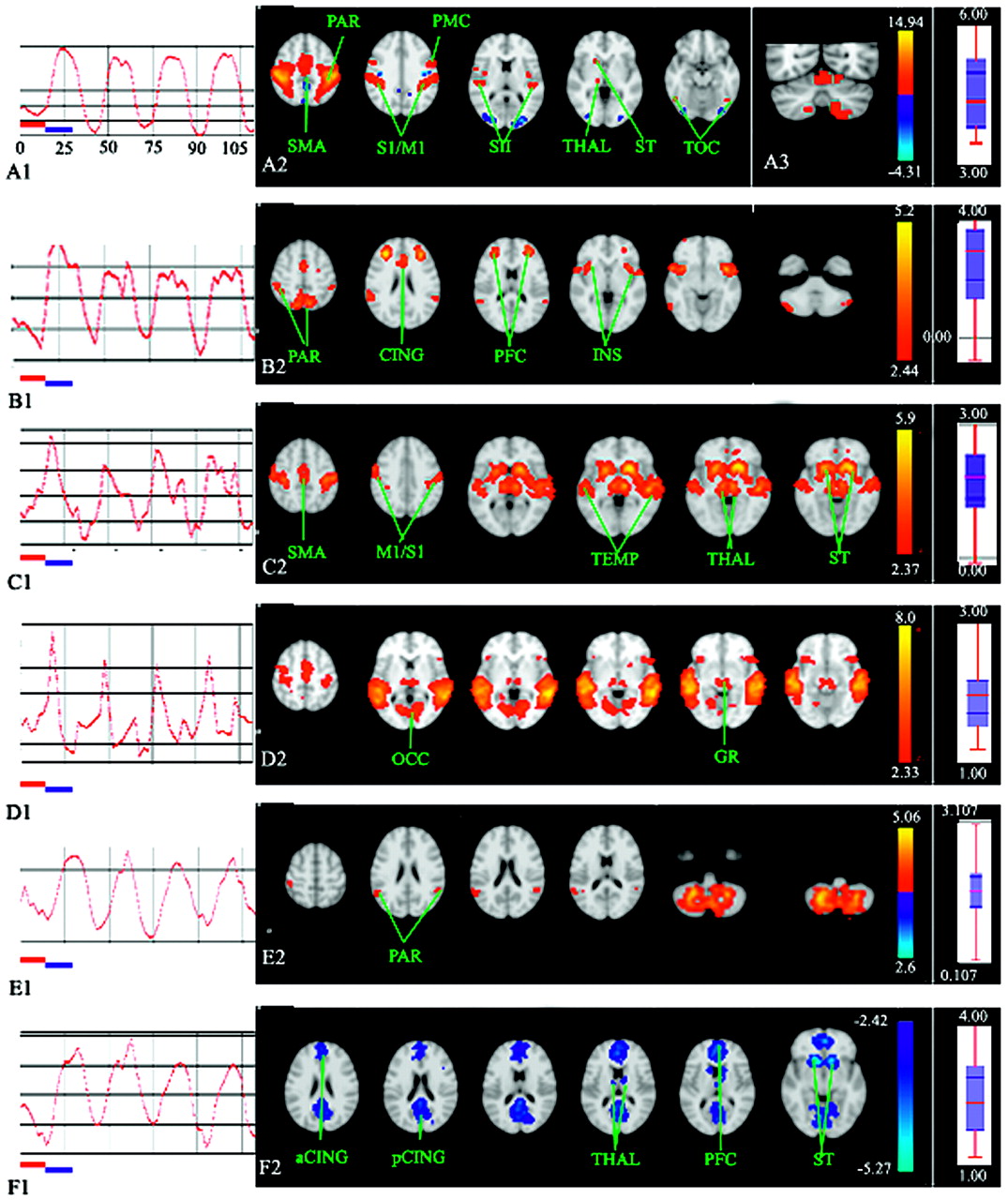
Bilateral activations were observed in Fig 1A1 and Table 1: the sensorimotor cortex, parietal operculum (secondary sensory area), supplementary motor area, and cerebellum (anterior lobe). The corresponding time course was strongly correlated with the succession of rest phases and activity phases (Fig 1A2).

A–B, For the motor task, multisubject (n = 14) spatial maps (A1–B1, axial sections) and time courses (A2–B2) of the independent components computed during the nondiscrimination task. A2–B2, Horizontal red and blue bars represent the first rest and activity phases, each lasting 15 seconds. A2–B2, Axial sections. Z-score is represented by a color gradient (vertical colored bar). Boxplots (on the extreme right side of the figures) display the relative response amplitude across the multisubject domain for the corresponding independent component sorted in decreasing order of the median response (median and upper and lower quartile are shown). CER indicates cerebellum; CING, cingulate cortex; GR, geniculate region; INS, insula; M1/S1, sensorimotor cortex; OCC, occipital cortex; PAR, parietal cortex; PFC, prefrontal cortex; PMC, premotor cortex; SII, secondary sensory area; SMA, supplementary motor area; ST, striatum; TEMP, temporal cortex; THAL, thalamus; TOC, temporo-occipital cortex; a, anterior; p, posterior.
Cluster localizations during the motor task for IC1 and IC2
Component 2: Sensorimotor Network.
Bilateral activations were observed in Fig 1B2 and Table 1: sensorimotor cortex, secondary sensory area, lateral premotor cortex, supplementary motor area, superior and inferior parietal lobules, insula, and prefrontal cortex. The left putamen was also activated. Bilateral putaminal activation was observed for a lower P threshold (>.5). Negative Z-values regarded as relative deactivation were located bilaterally in the prefrontal cortex, parietal cortex (precuneus), posterior cingulate cortex, occipital lobe, and parahippocampus. This deactivated circuit is partly reminiscent of the dorsal attention system devoted to top-down orienting of attention6 observed in the brain resting state.
Tactile Discrimination Task
Task-Related Components
The time courses of the 3 selected components were strongly correlated with the succession of rest phases and activity phases (Fig 2A1, -B1, -E1). Although not described here, these components were also present during the motor task.

Multisubject (n = 14) time-courses (A1–F1) and spatial maps (A2–F2, A3) of the independent components (A–F) computed during the tactile discrimination task. A1–F1, Horizontal red and blue bars represent the first rest and activity phases, each lasting 15 seconds. A2–F2, Axial sections. A3, Coronal section passing through the cerebellum. Z-score is represented by a color gradient (vertical colored bar). Boxplots (on the extreme right side of the figures) display the relative response amplitude across the multisubject domain for the corresponding independent component sorted in decreasing order of the median response (median and upper and lower quartile are shown). CER indicates cerebellum; CING, cingulate cortex; GR, geniculate region; INS, insula; M1/S1, sensorimotor cortex; OCC, occipital cortex; PAR, parietal cortex; PFC, prefrontal cortex; PMC, premotor cortex; SII, secondary sensory area; SMA, supplementary motor area; ST, striatum; TEMP, temporal cortex; THAL, thalamus; TOC, temporo-occipital cortex; a, anterior; p, posterior.
Component 1: Sensorimotor Network.
Bilateral activations were observed in Fig 2A2 and Table 2: sensorimotor cortex, lateral premotor cortex, supplementary motor area, superior and inferior parietal lobules, secondary sensory area, lateral occipital cortex (LOC), cingulate cortex, and cerebellum (anterior and posterior lobes, Fig 2A3). The right striatum was also activated. Left basal ganglionic activation was observed with P > .5. This sensorimotor network, though more strongly activated, was anatomically quite similar to the previous IC2 network characteristic of the motor task.
Cluster localizations during the tactile discrimination task for IC1 and IC2
Component 2: Executive Network.2
Bilateral activations were observed in Fig 2B2 and Table 2: sensorimotor cortex, lateral premotor cortex, supplementary motor area, pre-supplementary motor area, superior and inferior parietal lobules, secondary sensory area, insula, LOC, inferior prefrontal cortex, and prefrontal cortex. Activation was also noted in the right anterior cingulate cortex. Deactivation was present in superior and medial temporal, occipital and medial pericentral, and posterior cingulate cortices.
Transiently Task-Related Components
The time courses of the selected components were transiently synchronized with the beginning and/or end of the task (Fig 2C1–D1).
Component 4: Audiomotor Network.
Bilateral activations were observed in Fig 2C2: 1) the auditory system: superior temporal gyrus, Heschl gyrus, and language areas on the left side (planum temporale); and 2) the motor system: sensorimotor cortex, premotor cortex, supplementary motor area, striatum, and thalami.
Component 6: Audiovisuomotor Network.
This network shares the neural relays of the previous IC4 network. However, strong and specific activations were recorded bilaterally in the visual system (Fig 2D2): occipital lobe, mesencephalodiencephalic junction, probably the lateral geniculate area (x = 6 / −14, y = −34, z = −12), and the frontal eye fields.
Resting-State Network
This network was deactivated (Z < 0) during the discrimination phases (Fig 2F1) and was activated during the resting state.
Discussion
In this study, 3 concomitant neural networks specifically engaged during tactile-tactile matching discrimination were precisely differentiated and characterized; 1 of these networks was identified as being an executive network, previously described during the brain resting state; a differential involvement of the cerebellum and participation of a same given area in different networks were demonstrated; and transiently task-related and resting-state networks were identified.
First, the motor task recruited 2 circuits. The first motor circuit associated with IC1 corresponded to the circuit previously described during bimanual movements, in phase-index finger-thumb opposition,8,9 and included the sensorimotor cortex (motor execution), supplementary motor area (bimanual coordination, movement sequencing), and anterior cerebellar lobe (automation, movement correction). This circuit, involved in rhythmic and automated movements, therefore appeared to be responsible for movements required to pick up and drop objects. The second circuit displayed a broader anatomic extension and comprised new sensory areas, such as the parietal lobe and LOC. This circuit may be specifically activated when subjects grasped and held the chess pieces and, consequently, may underlie the adjustment of finger movements necessary to scale grip force according to proprioceptive and exteroceptive feedback. The parietal operculum and supramarginal gyrus have been shown to be involved in precision lifting tasks, especially during weight changes,10 in conjunction with the motor and premotor cortices11 and the basal ganglia.12 However, the most anterior activation of the parietal lobe within the intraparietal sulcus may process sensorimotor integration of finely tuned finger movements.13
Second, this sensorimotor network was also recruited during the tactile discrimination task, while the first motor network was no longer activated, implying that this sensorimotor network controls precisely tuned fingertip movements involved in palpation. These movements may be guided by sensory feedback (via the insula), mental representation of the manipulated objects (LOC), and sensory anticipation (at least, the parietal cortex).1 However, 3 differences in relation to the motor task were observed in this network: 1) stronger activation of the neural relay, probably caused by the greater attentional and computational load of the discrimination task; 2) activation of the cerebellar posterior lobe and anterior cingulate cortex, and 3) detection of no prefrontal activations. Therefore, only the cerebellar and cingulate involvement seems to be specific for the increased motor complexity of the task.
The posterior cerebellar lobe, especially the second homunculus (Lobule VIII), is involved with the supplementary motor area, in timing bimanual antiphase movements,8,9 and could therefore contribute to coordinating exploratory bilateral finger movements. The anterior cingulate cortex is involved in monitoring on-line performance, error detection, attention to action, suppression of inappropriate responses, and working memory.14,15 It can, therefore, be postulated that the participation of the sensorimotor network in more complex movements is mainly and nonspecifically under the dependence of the posterior cerebellum for motor coordination and the anterior cingulate cortex for executive control and motor readjustments when false anticipations or misidentifications of objects have occurred, for example.
During the tactile discrimination task, a second spatiotemporal process was also associated with IC2, referred to as the executive network. This network broadly encompassed 3 subcomponents: 1) fronto-cingulo-insular areas, 2) parietal areas, and 3) medial wall premotor areas (supplementary motor area, pre-supplementary motor area). The executive network, previously described during brain resting state, is centered on the ventro- and dorsolateral prefrontal cortices and anterior cingulate and paracingulate cortices and sends bias signals to modulate other areas. In this study, these modulated areas may include the premotor and parietal cortices.16 More precisely, there is an anatomofunctional parcellation of the prefrontal cortex partly related to haptic information: Brodman area (BA) 9/10 for general executive control, BA 10 for haptic working memory,17 BA 11 in attending tactile stimuli,18 and BA 9/46 for working memory of several pieces of information.19 These prefrontal executive networks would, therefore, participate in mental representation and comparison of objects in conjunction with LOC (3D haptic representation20) and the parietal cortex via the insula.
Mental representation and comparison are progressively performed through sensory inputs gathered by exploratory finger movements, which explains why supplementary motor area and pre-supplementary motor area activations were observed in the executive network. The supplementary motor area participates in complex bimanual motor timing and sequencing, whereas pre-supplementary motor area is connected with the prefrontal cortex; and the supplementary motor area and is involved in intentional task initiation and switching.21 The supplementary motor area and pre-supplementary motor area may, therefore, underlie volitional,22 motor planning and control, and correcting movements, whenever false identifications are detected by the anterior cingulate cortex on the basis of sensory feedback and sensory anticipations, respectively.
Finally, a third cerebelloparietal network was also strongly correlated with the discrimination task. Cerebellar activations were situated in the following areas: 1) the posterior lobe involved in bimanual coordination, on-line motor adjustments when unpredictable sensory stimuli occur,23 and response reassignment24; and 2) the dentate nuclei, the cerebellar output channel. The parietal node of this third network is located in the parietal lobe around the intraparietal sulcus, which plays a role in movement adaptation. Because the cerebellum is thought to predict the sensory consequences of motor commands via efferent copy,25 the function of this network could rely on rapid on-line motor adjustments based on mismatches between sensory anticipations and sensory feedback. In support of this hypothesis, the cerebellum has been implicated in conflict resolution,26 and the cerebellum is also reciprocally connected with the parietal cortex27,28 so that this cerebelloparietal network would constitute a third possible link between the sensorimotor and executive networks. In conclusion, TICA allowed discrimination of a widespread neural network involved in haptic representation into distinct specialized subnetworks possibly interconnected by common neural nodes.
Third, 2 networks anchored in the temporal lobe (IC4 and IC6), including the auditory areas, were activated at the beginning and, to a lesser extent, at the end of the activity phases (IC4). Their very transient activation, therefore, coincided with the auditory “go” and “stop” signals pronounced by the investigator. Because the IC4 network comprises motor and premotor areas and basal ganglia (action selection and inhibition of inappropriate movements), it could potentially participate in initiating and disengaging sensorimotor activation, possibly by exerting a positive modulatory influence, in conjunction with prefrontal and medial wall areas involved in the volitional control of movement.22 The second network (IC4) also showed activations located in the medial occipital cortex likely subserving mental imagery. However, the tactile discrimination task was accompanied by occipital deactivation, indicating that mental representation of chess pieces was related to LOC but not to primary and secondary visual areas.
Fourth, deactivation of the default-mode network dedicated to “stream of consciousness” and episodic memory4 was observed during tactile discrimination. This observation agrees with previous studies showing specific activation of this network during the resting state and anticorrelation with task-related areas4 (ie, engagement of the sensorimotor network is concomitantly accompanied by deactivation of the default-mode network). Strong deactivations also affected the caudate, accumbens, and thalamic nuclei, suggesting that this circuit may, at least partly, correspond to the limbic basal ganglionic loop.29
Fifth, some technical points and apparent limitations must be emphasized. We used a TR = 2500 ms, which can cause artifacts because of aliasing due to physiologic processes, and only 54 volumes were acquired though most studies applying TICA acquired at least 200 volumes, which can decrease the signal-intensity-to-noise ratio. Previous studies used a TR between 2000 and 3000 ms.2–6 Moreover, data-driven TICA is well suited to discriminate sources of noise, such as head motion, high-frequency noise, gradient instability, or field inhomogeneity, which give rise to known artifact patterns. Identification of relevant spatial maps was also guided by task-relation, statistical, and anatomic criteria to minimize operator dependency. Only spatial maps associated with 1) time-courses clearly (anti-) correlated with the design paradigm, 2) sufficiently explained variance (>5%), and 3) a neural circuit coherent with known anatomy and physiology were retained. For the resting state, only spatial maps associated with spontaneous low-frequency fluctuations (0.01–0.1 Hz) were retained.30 It may also be preferable to use more than 1 rater to select relevant maps, to ensure consensus interpretation of the results.
To maintain a good signal-intensity-to-noise ratio, we increased the posterior probability threshold to P > .7, though P is usually fixed at >.5.2 A high sampling level is required to estimate resting-state networks, resulting in a lower amplitude of the BOLD signal intensity than during stimulation tasks, and the time course of the signal intensity provides no information that can be used directly by the investigator to visually identify circuits. These precautions and the use of a high-field machine may have contributed to overcoming these technical limitations of this study, which demonstrates that short-acquisition sequences can be used for TICA investigation, an important point for future clinical applications, especially with noncompliant patients.
Conclusion
In conclusion, contrary to the GLM approach, TICA provides sharp insight into the dynamically evolving functional brain connectivity mobilized by the whole experimental paradigm. This multivariate model-free technique is able to differentiate activation of task-specific sequentially or concomitantly recruited networks and deactivation of task-unrelated or resting state networks. This study shows that the tactile-tactile matching discrimination task specifically activated 3 cooperative networks, including a sensorimotor network (motor program tk;4and execution), executive network (on-line elaboration and correction of haptic representation, working memory and sensory-guidance of exploratory movements), and a cerebelloparietal network (possibly sensory expectations) and deactivated the default-mode network and occipital areas and 2 transiently activated audiomotor and audiovisual circuits.
References
- Received April 8, 2008.
- Accepted after revision May 1, 2008.
- Copyright © American Society of Neuroradiology
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.