
Abstract
BACKGROUND AND PURPOSE: Gray matter heterotopia have been divided into three groups based on clinical and imaging characteristics: subependymal, subcortical, and band heterotopia. Nonetheless, subcortical heterotopia can have variable morphologic findings. The purpose of this study was to perform a morphologic analysis of a series of cases of subcortical heterotopia based on MR images, to correlate the morphologic appearance with clinical characteristics, and to speculate about the embryologic implications of our results.
METHODS: The MR imaging studies and clinical records of 24 patients with subcortical heterotopia were retrospectively reviewed. The morphologic findings of the heterotopia were recorded along with presence and type of associated malformations. These results were correlated with available data on development and neurologic status.
RESULTS: Analysis revealed that, in six cases, the heterotopia were composed exclusively of multiple nodules, in 13, they appeared primarily as curvilinear ribbons of cortex extending into the white matter, and in five, they had deep nodular regions with curvilinear areas more peripherally. All of the curvilinear regions were contiguous with the cerebral cortex in at least two locations. In eight cases, curvilinear heterotopia contained curvilinear areas of flow void that were thought to be blood vessels; in 10, they contained fluid resembling CSF. No difference in developmental or neurologic manifestations was noted among patients with heterotopia of different morphologic appearances.
CONCLUSION: Subcortical heterotopia can have nodular or curvilinear morphologic appearances. Although no difference was found in the clinical conditions of the patients with differing morphologic appearances, additional analysis of these patients or studies of animal models of these malformations may further our understanding of normal and abnormal brain development.
Gray matter heterotopia are fairly common malformations of cortical development that are thought to result from premature arrest of the migration of neurons from the germinal matrix in the wall of the lateral ventricle to the developing cerebral cortex (1, 2). Pathologists have traditionally classified heterotopia as laminar, nodular, leptomeningeal, or “double cortex” (also called band heterotopia) (2−4). Neuroradiologists, basing their ideas on MR imaging appearances, have classified the types of heterotopia as subependymal, subcortical, and band heterotopia (5, 6). In one article on MR imaging of heterotopia, a single case was mentioned in which the heterotopic gray matter seemed to represent an infolding of cortical gray matter (5). An initial review of the MR studies of patients with subcortical heterotopia at our institution revealed a number of patients with similar findings. Herein the clinical and radiologic findings of patients with curvilinear subcortical heterotopia are presented, the findings of these patients are compared with nodular subcortical heterotopia, and the nature of these malformations is explored.
Methods
A retrospective review of the radiology information system and teaching file films at University of California, San Francisco hospitals revealed MR imaging studies of 113 patients with heterotopia. Of these, 72 cases were classified as subependymal, 17 were classified as bands, and 24 were classified as subcortical. Further examination of the subcortical heterotopia MR findings revealed that in some cases, the heterotopia were composed primarily of multiple nodules but that others appeared primarily as ribbons of cortex extending into the white matter. These 24 studies are the subject of this report.
The 24 patients ranged in age from 3 days to 39 years, with a mean age of 17 years and a median age of 15 years. There were 13 female and 11 male patients. All available clinical histories and records of physical examinations were obtained. The MR studies had been performed using different imagers of different field strengths ranging from 0.3 to 1.5 T. As a result, the imaging parameters varied widely. All patients underwent at least one T1-weighted sequence and one T2-weighted sequence, and all underwent imaging in at least two planes. The images were carefully evaluated to analyze the morphologic appearance of the heterotopia (nodular versus curvilinear), the presence or absence of contiguity with the cortex, the presence of blood vessels within the heterotopia, and the presence of CSF within the heterotopia. The latter two features indicated contiguity with the subarachnoid space. All associated malformations, such as callosal agenesis, cerebellar malformations, variations in hemispheric size, dysplasia of the basal ganglia (defined as abnormalities in the shape or relative locations of the caudate nuclei, globi palladi, or putamina), and abnormal sulcation of the overlying cerebral cortex, were also noted.
Results
Clinical Data
In general, all of the patients had very similar clinical histories. Weakness, spasticity, and hyper-reflexia (long tract signs) were present on the body side contralateral to the side of the brain with heterotopia in 19 of the 24 patients. The two patients with bilateral heterotopia had bilateral long tract signs. Two patients with heterotopia restricted to the anterior portion of the frontal lobe had atrophy/muscle wasting in the arm contralateral to the malformation, but no long tract signs. The final patient, with a small occipital heterotopia, had normal results of the neurologic and physical examinations. No difference was detected in the clinical presentations of those with multinodular heterotopia as compared with those with convoluted curvilinear heterotopia.
All patients suffered from epilepsy. Initial seizures were detected as early as the first few days of life and as late as age 9 years. The mean age at seizure onset was 6 years. In the 16 patients for whom EEG results were available, slow wave activity, localized spikes, or both were detected in the area of the heterotopic gray matter.
Imaging Data
The MR images of all of the patients showed regions of gray matter within the cerebral hemispheric white matter. These varied in size from a few centimeters (Fig 1) to large regions (7−8 cm) that occupied much of a cerebral hemisphere (Figs 2 and 3). When heterotopia were unilateral, the affected part of the hemisphere was reduced in size compared with the normal contralateral hemisphere, with the reduction proportional to the total size of the heterotopia in all patients. This reduction seemed to be the result of a decreased volume of white matter in all patients. The overlying cortex appeared thinned, with abnormally shallow sulci, in all 24 patients (Figs 1–3).

FIG 1. Nodular subcortical heterotopia with callosal agenesis.
A, Axial spin-echo 3000/60 (TR/TE) image of a 1-month-old patient shows multiple nodular heterotopia (arrows) lining the frontal horn of the right lateral ventricle and extending into the center of the frontal white matter. The right hemisphere is reduced in size. The overlying cortex is thin and has a reduced number of sulci. The sulci present are abnormally shallow. The corpus callosum is agenetic.
B, Coronal spin-echo 600/16 image of the same patient shows the same findings.
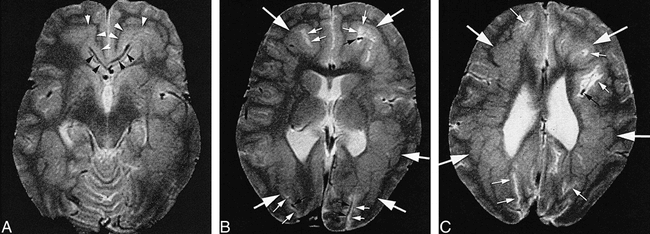
FIG 2. Curvilinear subcortical heterotopia.
A, Axial spin-echo (2500/80) image shows curvilinear subcortical heterotopia (white arrowheads) emanating from the pericallosal sulci bilaterally. Note the curvilinear flow voids (black arrowheads) of blood vessels extending into the heterotopia from the sulci.
B, Axial spin-echo (2500/80) image obtained at a higher level shows the heterotopia (large white arrows) extending all the way back to the occipital poles. Note the CSF (small white arrows) and the blood vessels (small black arrows) coursing within the heterotopia. The cerebral cortex is thin, and the sulci are shallow.
C, Axial spin-echo (2500/80) image obtained at a still higher level shows the contiguity of the curvilinear heterotopia (large white arrows) from the frontal to the occipital lobes. Note that CSF (small white arrows) and blood vessels (small black arrow) are present, even at the center of the heterotopia.
Six patients seemed to have purely nodular subcortical heterotopia, consisting of multiple nodules extending from the ventricular surface outward into the hemispheric white matter (Fig 1). No definite contiguity of the heterotopia with the cerebral cortex was detected. All six patients had anomalies of the corpus callosum (agenesis in three and hypogenesis in three). The basal ganglia were dysplastic on the side of the heterotopia in five of the six patients.
Thirteen patients had purely curvilinear subcortical heterotopia. Careful analysis revealed that, in each of these cases, heterotopia had the appearance of enfolded cortex in clear contiguity with the cortex in at least two locations (Fig 2). In eight patients, blood vessels could be seen coursing within the layers of gray matter (Fig 2). In 10 patients, fluid with the signal characteristics of CSF could be seen within the gray matter convolutions of the heterotopia (Fig 2). When both fluid and vessels were present, the vessels seemed to course within the fluid (Fig 2). Ten patients had associated anomalies (agenesis or hypogenesis) of the corpus callosum, whereas eight had dysplastic ipsilateral basal ganglia.
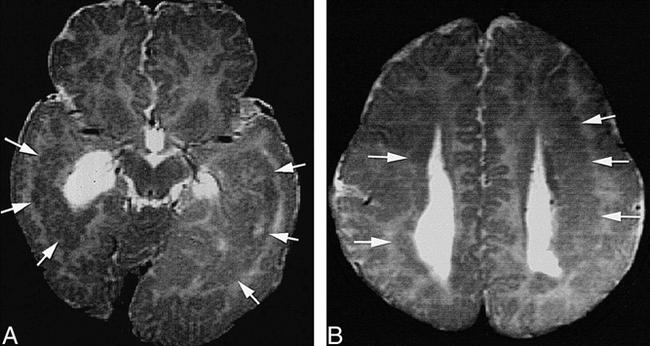
Five patients had regions of both nodular and curvilinear subcortical heterotopia (Fig 3). Isolated gray matter nodules were seen in the deep portions of the mass (closest to the ventricle) and curvilinear convolutions were seen in more superficial portions, with an almost imperceptible transition between them (Fig 3). The nodular regions of heterotopia were identical to the purely nodular heterotopia, whereas the convoluted portions were identical to the convoluted heterotopia. All of the five had anomalies of the corpus callosum (agenesis in two and hypogenesis in three). The basal ganglia were dysplastic on the side of the heterotopia in all five.

FIG 3. Mixed curvilinear/nodular subcortical heterotopia.
A, Axial spin-echo (2500/80) image shows nodular heterotopia (white arrows) in the periventricular white matter bilaterally. No cortical connection is seen at this level.
B, Axial spin-echo (2500/80) image obtained at a higher level shows that the heterotopia (white arrows) are contiguous with the cortex bilaterally.
Discussion
Heterotopia are common malformations that typically are associated with epilepsy and variable developmental delay in children and young adults (5, 7−9). Classification of heterotopia can serve two purposes; one clinical (to direct therapy and allow prognosis of individual patients) and one research-oriented (to help find mechanisms of formation that may ultimately allow therapeutic intervention). The classification of heterotopia into subependymal, subcortical, and band types has been considered to be useful because these three groups of patients have different clinical presentations and different underlying genetic disorders (5, 10−14). It was, therefore, worthwhile to investigate whether the differing morphologic appearances of subcortical heterotopia were of clinical significance.
This morphologic analysis of subcortical heterotopia indicates that such heterotopia have at least two different morphologic configurations, which may be called the subcortical nodular heterotopia and the subcortical curvilinear heterotopia. A third type, which we will call the mixed subcortical heterotopia, seems to be composed of a mixture of the first two. Considering that the clinical manifestations of the affected patients and the anomalies associated with the different morphologic configurations appear identical, this differentiation is unlikely to be of any clinical significance. It is possible, however, that analysis of these differing morphologic appearances may allow some insight into the mechanisms by which subcortical heterotopia develop; such insight may aid in our understanding of normal and abnormal brain development.
The most common type of heterotopic gray matter is the subependymal nodular type (2, 5). Subependymal heterotopia can be isolated or associated with other anomalies of the brain (15, 16). When heterotopia are isolated, patients typically have normal results of neurologic examinations and normal early development; they come to medical attention with the onset of epilepsy, usually during the second decade of life (5, 7). Heterotopia can be caused in animals by X-irradiation or by toxin administration (17), both of which presumably disrupt the normal process of neuronal migration. The process of neuronal migration is dependent on interaction of a receptor on the surface of the migrating neuroblast with a ligand on the adjacent radial glial cell. The activated receptor then initiates a series of intracellular reactions that result in extension of the neuronal growth cone along the radial glial cell. The cell body then follows the growth cone. Ultimately, the cell body of the neuroblast reaches the developing cortex, where it receives a signal to dissociate from the radial glial cell and become a part of the cortex (18−25). It is likely that most cases of nodular heterotopia are the result of the failure of one or more aspects of this process. Recent work has shown that some cases of subependymal heterotopia are X-linked, with the responsible gene, called filamin 1, localized to Xq28 (10). Male patients with X-linked subependymal heterotopia have widespread cerebral abnormalities and significantly worse clinical outcomes (26−28). Filamin 1 shows a high level of expression in the developing cortex and is required for migration of neuroblasts to the cortex. The gene product has been identified and seems to be an actin cross-linking phosphoprotein (11). After a receptor on the surface of the migrating neuron is activated by interaction with a ligand on the radial glia cell, this phosphoprotein induces actin reorganization. Actin reorganization is required for the linkage of growth cones to the actin cytoskeleton and is thus necessary for the locomotion of the migrating cell (11). Microscopically, subependymal heterotopia are composed primarily of immature neurons and astrocytes; the neurons (and thus the heterotopia) increase in size with increasing age (2). The neurons in these heterotopic nodules have a limited number of connections with each other and with the overlying cortex (29), which may also be dysplastic (30). This limited connectivity may explain why the heterotopia are sometimes, but not always, the source of seizures in affected patients (7, 31). This lack of connectivity has also been postulated to be the cause of the neuronal immaturity within heterotopia (neuronal complexity seems proportional to its connectivity) (29). Furthermore, the immaturity of inhibitory gamma aminobutyric acid neurons within these nodules is speculated to produce an excess of excitatory over inhibitory neuronal circuitry, resulting in the seizure activity that is so common in affected patients (29).
Most occurrences of band heterotopia seem to be X-linked; the responsible gene is called doublecortin or XLIS (12, 32). Most patients with band heterotopia have some degree of developmental delay, the severity of which seems to be related to the thickness of the band (3). Seizures are very common and often mixed in type (3, 33). The XLIS gene is located in chromosome Xq23.3−q24 and encodes a novel protein that is involved in an unknown intracellular signaling pathway (12, 32, 34). Chromosome rearrangements or point mutations of XLIS cause lissencephaly in affected male patients and band heterotopia in affected female patients (35). No correlation has been detected between genotype and phenotype, indicating that the variable severity of this disorder in female patients is most likely the result of variable X-inactivation in neuronal precursor cells (13).
Much less is known about subcortical heterotopia than about subependymal or band heterotopia. The concept of subcortical heterotopia is a radiologic one; similar regions are classified as either nodular heterotopia or laminar heterotopia in developmental neuropathology texts (2, 15). Few of our cases of subcortical heterotopia, however, were purely nodular nor did they fit the criteria for laminar heterotopia, which are defined by Friede (15) as “bilateral, symmetrical ribbons of gray matter… between the cortex and ventricular walls, separated from both by layers of white matter.” This definition of laminar heterotopia describes the features that are currently generally attributed to band heterotopia. Norman et al (2) separated laminar heterotopia from band heterotopia in their classification but noted that they had no personal experience with either. Most of the heterotopia in our series had an appearance that somewhat resembled convolutions of cortex extending into the white matter from the cortical surface (Fig 2). In several patients, blood vessels and fluid were seen within the heterotopic region (Fig 2), strongly suggesting communication with the subarachnoid space. How does this communication arise? It is difficult to answer this question if the subcortical heterotopia are the result of simple arrest of the radial migration of neurons from the germinal zone to the cortex. Another possible mechanism is suggested by observations of heterotopia formation in a tish rat model developed by Lee et al (36, 37). This rat develops subcortical heterotopia below the frontal and parietal cortices that are somewhat similar in appearance to those of human subcortical heterotopia (36) (although blood vessels and CSF are not described within the heterotopia of the tish rat). In contrast to the sparse connectivity of neurons in subependymal heterotopia, the tish heterotopia have normal afferent and efferent connections with subcortical structures such as the thalami (38). The authors note that the affected rats seem to develop a secondary (heterotopic) germinal zone in the intermediate zone of the developing brain. Moreover, they provide evidence that neurons migrate peripherally into the heterotopia from the normotopic germinal zone located at the ventricular wall and centrally into the heterotopia from the heterotopic germinal zone in the subcortical region. The authors provide evidence that neurons generated in the heterotopic germinal zone migrate both inward to the heterotopia and outward to the cortex. The presence of this heterotopic proliferative zone could possibly explain both the contiguity of the heterotopia with the cortex and the connection between the heterotopia and the subarachnoid space. If some neurons produced by the heterotopic proliferative zone persisted between the cortex and the heterotopia, it could act as a bridge of gray matter between the two. It is also possible that the same factors that induce pial capillaries to perforate the brain surface and grow into the cerebral cortex (39, 40) may be generated in the heterotopic cortex, inducing the capillaries to continue growing inward into the substance of the subcortical heterotopia. If the vessels grow into the heterotopia, it naturally follows that a distinct perivascular tissue space, called the Virchow-Robin space or Virchow-Robin compartment (39, 40) would form around it. The Virchow-Robin compartment is thought to have some communication with the subpial space and hence the CSF circulation (40). This communication would explain the fluid seen around the vessels that were identified within the curvilinear subcortical heterotopia. Although the nodular heterotopia seen deep to the curvilinear heterotopia in five patients are not explained by the postulate of a heterotopic germinal zone, it is not unreasonable to suggest that the normotopic germinal zone in patients with a heterotopic zone and the neuroblasts produced therein would have a higher than normal likelihood of being abnormal. These neuroblasts could have abnormalities that would cause premature termination of migration, producing nodular heterotopia. Alternatively, centrally migrating neuroblasts from the heterotopic germinal zone could terminate migration in the deep white matter, causing the same result. Further experiments with appropriate animal models would be necessary to determine which, if either, of these explanations is correct.
One final comment concerns the nomenclature used in describing heterotopia. It is the opinion of this author that the term laminar heterotopia is confusing. The way Friede (15) defines the term, it is synonymous with band heterotopia (also called double cortex); however, the example he shows looks more like multiple nodular heterotopia. Des Portes (13) refers to band heterotopia as subcortical laminar heterotopia. Radiologically, band heterotopia and subcortical heterotopia are distinct. Band heterotopia consist of smooth layers of gray matter that often follow the curvature of the overlying cortex. They are not convoluted nor are they contiguous with the overlying cortex. They do not contain blood vessels or CSF. Subcortical heterotopia usually consist of swirling, heterogeneous, curvilinear masses of gray matter often containing blood vessels and CSF. They are essentially always contiguous with the overlying cortex and the underlying ventricular surface. Pathologists, unfortunately, suffer from being able to cut brains in only a single plane. Thus, although they get a 3D view of the surface of the brain, they are somewhat limited in their ability to assess structures, such as heterotopia, that are situated in the white matter. In a single section, curvilinear subcortical heterotopia will often look like multiple small nodules. Multiple images in multiple planes are required to understand the complex curvilinear structure of the region fully. The example presented by Friede (15), which is described as a nodular heterotopia, would likely be classified as curvilinear subcortical heterotopia if imaged in multiple planes at multiple levels. Why is it important to make this differentiation? The discussion herein should make that clear. Nodular heterotopia are likely the result of either lack of migration or of simple premature termination of migration, whereas curvilinear subcortical heterotopia are likely the result of a much more complex process. Some might consider curvilinear subcortical heterotopia to be infoldings of dysplastic cortex and not true heterotopia. I suggest that in a process as complex as cortical development, the name given to these malformations are less important than an understanding of their clinical manifestations and the mechanisms by which they develop. By learning and understanding the differences in the embryogeneses of these cortical malformations, we will come closer to understanding the process of brain development.
Conclusion
This morphologic analysis of the MR studies of 24 patients with subcortical heterotopia suggests that at least two different types of subcortical heterotopia exist, a nodular form and a curvilinear form that seems to communicate with the overlying cerebral cortex and subarachnoid space. Several patients with a mixed form were also identified. No differences in the clinical manifestations or in the associated anomalies were identified among these three groups of patients. Correlation with animal models of heterotopia suggest possible mechanisms by which these heterotopia form.
Footnotes
↵1 Address reprint requests to A. James Barkovich, MD, Department of Radiology, Neuroradiology Section L 371, University of California, San Francisco, 505 Parnassus Avenue, San Francisco, CA 94143-0628.
References
- Received June 18, 1999.
- Copyright © American Society of Neuroradiology
In this issue




{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Subcortical heterotopic gray matter brain malformations: Classification study of 107 individuals
- Malformations of Cortical Development and Epilepsy
- Ultra-High-Field MR Imaging in Polymicrogyria and Epilepsy
- Disorders of Cortical Formation: MR Imaging Features
- Benefits, risks and ethical considerations in translation of stem cell research to clinical applications in Parkinson's disease
- Gray matter heterotopia