
Abstract
BACKGROUND AND PURPOSE: Repetitive motor tasks that produce sustained neuronal activity in the sensorimotor cortex produce transient neuronal activity in subcortical regions. We tested the hypothesis that a reference function modeling a transient hemodynamic response would more reliably detect activation in the basal ganglia than would a conventional reference function, which models a sustained hemodynamic response.
METHODS: Functional MR imaging data were acquired in eight subjects performing an alternating-hand finger-tapping task. Postprocessing was performed by cross-correlation to two types of reference functions: one that models a sustained hemodynamic response to finger tapping and one that models an initial transient hemodynamic response. Activation in the sensorimotor cortex, supplementary motor area, cerebellum, thalamus, and corpus striatum was tabulated for each reference function.
RESULTS: With the conventional boxcar reference function, activation was detected in the sensorimotor cortex, supplementary motor area, and cerebellum, but intermittently in the corpus striatum in all subjects. With the reference function for a transient response, activation in the corpus striatum was not detected in all subjects.
CONCLUSION: In the corpus striatum, activation is detected more frequently with a reference function that models a transient response. Activated cortical and subcortical regions can be mapped with an alternating-hand finger-tapping paradigm and a combination of reference functions.
As demonstrated with functional MR imaging, the finger-tapping paradigm regularly produces activation in the sensorimotor cortex (SMC) (1) but not regularly in the subcortical brain structures, which are also known to be involved in the production of voluntary movements. Functional MR imaging activation represents a change in signal intensity caused by regional blood oxygen level–dependent (BOLD) contrast changes corresponding to finger-tapping tasks (2). These tasks have been used extensively in studies of patients who are scheduled for neurosurgical procedures (3) and in healthy subjects to study the organization of cognitive functions (4). Activation in the basal ganglia associated with performance of motor tasks has been reported inconsistently (5–9).
The amount and the location of activation detected during performance of a motor task depend on the task paradigm and on the regional hemodynamic response. The hemodynamic response to a motor task is more sustained in the SMC and is more transient in the dorsal prefrontal and medial frontal cortices (7). In the performance of repetitive motor tasks, basal ganglia circuits have been implicated in the selection and initiation of motor task performance (10–12). Therefore, we tested the hypothesis that the hemodynamic response to finger tapping may have a shorter duration in the basal ganglia than in the SMC. We acquired functional MR imaging data sets in subjects performing alternate hand- and finger-tapping tasks and analyzed the data with reference functions that modeled a prolonged hemodynamic response and with others that modeled a transient response.
Methods
Eight right-handed volunteers (four men and four women) with no significant medical history or current medical problems were enrolled after they signed a consent in accordance with institutional policy. Handedness was determined by the Edinburgh inventory multiresponse questionnaire (13). Before the scanning session, the task was described by the investigator and briefly practiced by each subject to confirm his or her comprehension and ability to perform the task. The task paradigm included four cycles (of 40 seconds' duration) of self-paced finger tapping alternating with rest periods (of 20 seconds' duration) as cued by the investigator, after an initial rest period of 28 seconds. For the finger-tapping task, the subject was instructed to oppose each finger with the thumb in sequence. During each finger-tapping period the subject was cued first to use the right hand for 20 seconds and then to use the left hand for 20 seconds. Subjects were asked to perform the finger tapping as rapidly as they could, and also to try to tap the fingers of each hand at the same rate. Task performance was visually monitored during the functional MR imaging studies. Subjects were instructed to keep their eyes closed throughout the imaging study, to concentrate on tapping as quickly and evenly as possible when given auditory cues, to avoid head movement, and to refrain as much as possible from higher cognitive processes during the rest periods.
Imaging was performed with a clinical 1.5-T magnet equipped with high-speed gradients for whole-body echo-planar imaging (EPI). The subject's head was positioned within a prototype radio-frequency quadrature bird cage coil with foam padding to provide comfort and to minimize head movements. Aircraft-type earphones with additional foam padding were placed in the external auditory canals to reduce the subject's exposure to ambient scanner noise and to enable communication. Preliminary anatomic images included a sagittal localizer image followed by a 3D spoiled gradient-recalled imaging whole-brain volume with the following parameters: 21/7 (TR/TE), 40° flip angle, 24-cm field of view (FOV), 256 × 256 matrix, 124 contiguous axial slices (including the vertex) through the cerebellum, and a 1.2-mm slice thickness. A series of 18 coronal T1-weighted spin-echo images was acquired from the occipital pole to the anterior frontal lobe with parameters of 500/8, 24-cm FOV, 256 × 192 matrix, and a 7-mm slice thickness with a 2-mm gap.
EPI functional MR imaging studies were acquired at the same slice locations and with the same thickness and gap as the spin-echo coronal anatomic series. EPI parameters included gradient-recalled echo single-shot acquisition with 2000/40, 90° flip angle, 24-cm FOV, 64 × 64 matrix (in-plane resolution of 3.75 × 3.75 mm), and ±62.5 kHz receiver bandwidth. Shimming was optimized with an automated gradient-shim sequence on a manually selected 3D brain volume region of interest (ROI). Transmit gain and resonant frequency were also manually tuned before the functional studies were obtained. Duration of the functional MR imaging EPI scan was 4 minutes 28 seconds (134 data time points).
Analysis
EPI raw data were filtered in the spatial frequency domain using a Hamming low-pass filter (14) then reconstructed into individual slice-location time courses. These reconstructed time course files were then checked for head motion and realigned using a 3D spatial registration algorithm (AFNI; Bob Cox, Medical College of Wisconsin). All scans with motion exceeding one voxel were excluded. The time course plots in cortical and subcortical regions with possible motor activation were inspected to determine the temporal nature of the hemodynamic response.
Correlation between each time course plot and the reference function was determined by means of Student's t test. Each time course plot was fitted to reference functions with a generalized least-squares fitting algorithm (15). The fitted functions included a constant (baseline signal level), a ramp (to allow for possible linear signal drift), and a reference function modeling the presumed stimulus responses. Five different reference functions were used (Fig 1), including two conventional functions that modeled a sustained hemodynamic response (reference functions 1 and 2), two functions that modeled a transient hemodynamic response (reference functions 3 and 4), and one function designed to detect the activation when either or both hands were active (reference function 5). Reference functions 1 and 2 were boxcars of unit amplitude for 20 seconds and zero amplitude for 40 seconds. For reference function 1, the departure from baseline corresponded to the 20 seconds of right-handed finger tapping, and for reference function 2 it corresponded to the left-handed finger tapping. Reference functions 3 and 4, designed to model a hypothesized transient hemodynamic response at the initiation of each task cycle, were boxcars with unit amplitude for 2 seconds and zero amplitude for 58 seconds. For reference function 3, the departure from baseline coincided with the first 2 seconds (first TR) of the right-handed task, and for reference function 4 it corresponded to the first 2 seconds (first TR) of the left-handed finger task. Reference function 5 had a boxcar, with unit amplitude for the 40-second period during which either hand was active and zero amplitude during rest. All boxcars were convolved with a Poisson function, with a mean of 6 seconds to adjust for the expected delay of the hemodynamic response to neuronal activity. Pixels with a t statistic exceeding a threshold value of P < .001 were mapped onto the anatomic images as regions of activation. Activation in the SMC, supplementary motor area (SMA), superior cerebellum, inferior cerebellum, thalamus, caudate nucleus, putamen, and globus pallidus was identified by conventional parcellation methods (16) and tabulated for each reference function.

Reference functions and task timing
Time courses from voxels within cortical and subcortical regions were inspected. For selected voxels, the signal intensity during each of the four 60-second epochs was averaged using the AFNI single trial averaging plug-in. These time course graphs represent the average across each of the four data points that occurs during the same temporal location within each repetitive hand task. The duration of the hemodynamic response was compared for cortical and subcortical regions.
Results
All subjects complied with the task paradigm, and technically satisfactory functional MR images, without significant motion artifacts, were obtained for each subject. Results for reference functions 1 and 2 are listed in Table 1. The reference function for prolonged response to the right-handed task (reference function 1) detected activation in the left SMC for all eight subjects, and the reference function for prolonged response to the left-handed task (reference function 2) detected activation in the right SMC in each subject (Fig 2). For reference functions 1 and 2, ipsilateral SMC activation was identified in four and seven subjects, respectively. The contralateral SMA was activated in seven subjects for the right-handed reference function (reference function 1). In eight subjects the SMA was activated bilaterally with the left-handed task function (reference function 2). Ipsilateral superior cerebellar activation was seen in all subjects, with contralateral response seen in seven subjects for reference function 1. Ipsilateral and contralateral activation was seen in eight subjects for reference function 2. The ipsilateral inferior cerebellum was also activated for all subjects, with contralateral response in two subjects for the right-handed function (reference function 1) and in six subjects for the left-handed function (reference function 2). Seven subjects had activation in the contralateral thalamus for reference functions 1 and 2. Two subjects had activation in the ipsilateral thalamus for reference function 1 or 2. In the striatum, activation was seen inconsistently in either the ipsilateral or contralateral hemisphere. No activation was seen in the globus pallidus for reference functions 1 or 2.
Number of subjects who showed activation in specific regions for reference functions 1 and 2

A–F, Functional MR images comparing activation detected by reference function 1 (right-handed task) (A–C) and reference function 2 (left-handed task) (D–F). Activation is detected in the SMC, thalamus, putamen, and cerebellum; activation is contralateral to the fingers that were active in the SMC and the thalamus, bilateral in the putamen, and ipsilateral in the cerebellum. (Note that all images are conventional radiologic display format, with the subject's right side appearing on the viewer's left.)
Reference functions 3 and 4, corresponding respectively to the first 2 seconds of right-handed (reference function 3) and left-handed (reference function 4) finger tapping, showed a decrease in sensitivity to activation in the SMC, SMA, thalamus, and cerebellum as compared with reference functions 1 and 2. Differences were noted for the striatum (Table 2). For the ipsilateral or contralateral striatum, reference functions 3 and 4 showed activation more consistently than did reference functions 1 or 2 (Fig 3). Globus pallidus activation was identified in two subjects, and only with reference functions 3 and 4. Activation in the auditory cortex was also identified in most subjects with reference functions 3 and 4 coincidentally with the auditory cues given to the subjects for task timing at the beginning of each task epoch.
Number of subjects who showed activation in the basal ganglia for reference functions 1–4

A and B, Functional MR images showing subcortical activation for the same subject as in figure 2 detected by reference function 3 (right-handed initiation) (A) and reference function 4 (left-handed initiation) (B). Activation is detected in the putamen, thalamus, and transverse temporal gyrus. As compared with reference functions 1 and 2 (see fig 2B and E), more activation is detected in the putamen
The fifth reference function, corresponding to the 40 seconds in which left-handed and then right-handed finger tapping was performed, identified activation in the SMC, SMA, and cerebellum in all subjects. In the time course plots of activation, the SMA in particular exhibited a high correspondence to this reference function. It identified thalamic activation in five subjects and striatum activation in five subjects. Activation related to the right or the left hand could not be distinguished on the images. Compared with reference functions 1 and 2, reference function 5 had poorer spatial correlation with activation of the pre- and postcentral gyri (Fig 4). This result was to be expected, as the 40-second reference function was not specific to the 20-second task cycle for each hand but included 20 seconds when either of the hands was also inactive.

Functional MR images showing activation detected by reference function 5. Activation is detected in the SMC, SMA, thalamus, and cerebellum. As compared with the other reference functions (figs 2 and 3), activation is detected bilaterally in the SMC, thalamus, and cerebellum, but with decreased specificity for the SMC
The time course of the BOLD effect differed between the cortical and subcortical regions (Fig 5). Time courses from the SMC and superior cerebellum had more sustained responses to finger tapping than did those in the basal ganglia. The temporal changes in the cortex corresponded better to reference functions 1 and 2; time courses from the putamen corresponded better to reference functions 3 or 4 than to reference functions 1 or 2 (Fig 5F). The averaged time course for activation in the putamen was markedly shorter than was the averaged time course for activation in the SMC (Fig 6). In both the right and left putamen, an increase in signal intensity corresponding to the initiation of left- and right-handed finger tapping was evident. The time course plots for the SMA showed a sustained signal increase that corresponded well to reference function 5 (Fig 5E). In all brain regions, maximal hemodynamic response was delayed by about 6 seconds with respect to the initiation of the task (as measured by a 2-second TR).
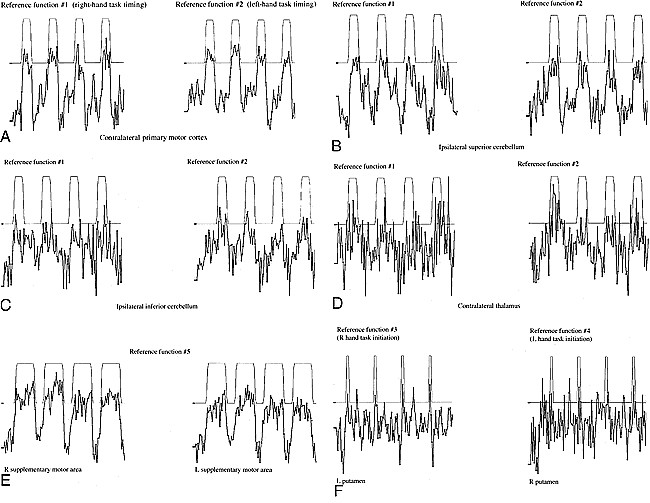
Representative time courses from selected ROIs.
A, Right and left primary motor cortex.
B, Right and left superior cerebellum.
C, Right and left inferior cerebellum.
D, Right and left thalamus.
E, Right and left SMA.
F, Right and left putamen.
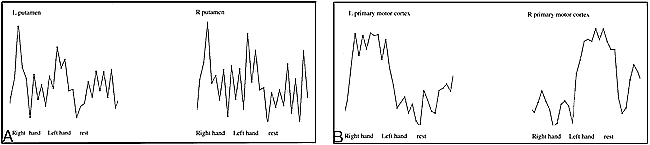
A and B, Time courses are averaged across the four epochs from representative pixels in the right and left putamen (A) and left and right primary motor cortex (B). Vertical gray lines indicate timing intervals for the initiation of each 20-second cycle of right- and left-handed activation and rest
Discussion
This study shows that reference functions that model a prolonged or a transient response differ in their sensitivity to activation in the basal ganglia associated with a finger-tapping task. Detection of activation in the basal ganglia and specifically in the globus pallidus and striatum was improved by the use of reference functions that corresponded temporally to the transient hemodynamic response in the basal ganglia at the initiation of a repetitive motor task. The third and fourth reference functions showed basal ganglia activation in seven of eight subjects, whereas the first and second reference functions showed basal ganglia activation in five subjects. Time course plots confirmed the temporal differences between the hemodynamic responses in the basal ganglia and the SMC associated with the finger-tapping task. The hemodynamic responses were more prolonged in the SMC than in the basal ganglia. A standard cross-correlation technique (17) applied to the finger-tapping task does not show basal ganglion activation as effectively as does a method that assumes a shorter response. None of the reference functions invariably showed activation within the basal ganglia or thalamus.
The sensitivity and specificity of the reference function to activation in different parts of the motor systems depend on many factors, including the temporal length of the function. Cross-correlation analysis with different reference functions can result in commonality of detected activations (Table 2), especially when the reference functions are not mutually exclusive. In this study, the first and second pairs of reference functions shared a partial temporal overlap corresponding to the initial 2 seconds of each task cycle. Similarly, the fifth reference function overlapped with the others (Fig 1). Reference functions of short duration, such as reference functions 3 and 4, were more sensitive to slight variations in response timing. The commonality of activations shown in Table 2 also indicates subject variability for hemodynamic responses among the idealized references. Higher statistical power through averaging across a greater number of task cycles (more than the four cycles in this motor task paradigm) might contribute to a greater specificity among the results for each reference function. The tabulated trend toward increased activation in the basal ganglia with the transient reference functions, combined with the averaged time course analysis and activation maps, indicates an improved sensitivity to motor task activation in the basal ganglia that is characterized by a transient initial hemodynamic response.
In previous functional MR imaging studies, investigators have identified basal ganglia activation associated with finger tapping and other motor tasks. Reichenbach et al (8) tabulated activation in cortical structures as well as in the putamen and cerebellum in subjects performing a thumb-flexion task paradigm. While exercising their dominant hand, all seven of these subjects activated the contralateral putamen and ipsilateral cerebellum, and five subjects activated the ipsilateral putamen. In that study, as in ours, bilateral activation occurred more often with the left-handed task than with the right-handed task. However, Reichenbach et al did not report activation in the thalamus as frequently as it occurred in our study, perhaps owing to their use of a simple thumb-flexion task. Moriyama et al (9) reported contralateral thalamic activation in two of five subjects in response to a complex finger-opposition task. They observed contralateral cortical and ipsilateral cerebellar activation and no activation in either the putamen or globus pallidus. Samuel et al (7) found activation in the contralateral putamen from data averaged across five subjects performing a joystick manipulation task. In two other functional MR imaging studies, one at 3.0 T (6) and one with a fast low-angle shot sequence at 1.5 T (5), basal ganglia activation was identified in conjunction with motor tasks. Lehericy et al (6) used a simple thumb or toe flexion/extension task, and Bucher et al (5) used hand supination/pronation. In both these studies, activation seen in the putamen was predominantly contralateral. Lehericy et al (6) also found contralateral thalamic activation in nine of 10 subjects for thumb movement measured at 3.0 T.
Investigators using positron emission tomography (PET) have consistently found regional cerebral blood flow (rCBF) increases in the basal ganglia associated with the performance of various motor functions (18). PET (12, 19) and in vivo physiological studies (10, 11) have implicated the basal ganglia in the functionally connected loop for motor task learning, initiation, and execution. In PET studies of healthy subjects performing finger-opposition task paradigms, rCBF increases were seen in the contralateral SMC and lentiform nucleus, with primarily ipsilateral increases in the cerebellum (18, 20). Our study demonstrates the possibility of a functional MR imaging hemodynamic corollary to the neuronal and PET findings.
We did not use a variety of task paradigms in this study, although previous reports have indicated that tasks of different complexity produce different activation patterns (4, 21, 22). The alternating finger-tapping paradigm was used to minimize artifacts in the midline veins and to encode laterality of activation (23). A study of similar alternating-hand tasks of different complexities, analyzed with reference functions relevant to the performance aspects of each task, might further define the functional MR imaging signal responses to motor activation.
Intrasubject reproducibility was not tested as part of this study. The test-retest repeatability of cortical motor activation has been studied (21, 24, 25). Scholz et al (21) reported high intertask variability for activations within the basal ganglia. Some of the variability may be due to the diminished sensitivity of the conventional reference function for transient activation. Other factors include the relatively low signal intensity increase during activation and the localized dephasing effects due to regional iron deposition. Some of the variance of basal ganglia activation in this study may also have been due to these effects.
Conclusion
Different patterns of functional MR imaging motor task activation are detected as the temporal duration of the reference function is changed. A reference function that models a transient hemodynamic response detects activation in the basal ganglia more frequently than does the conventional reference function that models a sustained hemodynamic response. Functional MR imaging hemodynamic activation responses secondary to an alternating-hand finger-tapping task can be mapped to cortical and subcortical regions using cross-correlation analysis with a combination of different reference functions.
Footnotes
↵1 Address reprint requests to Chad H. Moritz, Department of Radiology, Clinical Sciences Center, Module E3, 600 Highland Ave, Madison, WI 53792.
References
- Received August 6, 1999.
- Accepted after revision January 20, 2000.
- Copyright © American Society of Neuroradiology




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}