
Abstract
BACKGROUND AND PURPOSE: Anosmias with chromosomal disorders has been well investigated. However, isolated anosmia (IA) has received less attention, although it occurs more frequently. We compared frontobasal structures in patients with IA since birth or early childhood with those in control subjects.
METHODS: Imaging findings obtained in 16 patients with IA were compared with those obtained in eight control subjects. Imaging was performed with a standard quadrature head coil at 1.5 T. T1-weighted spin-echo (coronal plane perpendicular to frontal skull base; section thickness, 3 mm; pixels, 0.43 × 0.39 mm) and sagittal T1-weighted magnetization-prepared rapid gradient-echo (voxels, 1.0 × 1.0 × 1.0 mm) sequences were performed. We assessed the length and depth of the olfactory sulcus, olfactory bulb volume, and olfactory sulcus depth in the plane of the posterior tangent through the eyeballs (PPTE).
RESULTS: Five patients with IA had bilateral hypoplastic olfactory bulbs. Three patients with IA had hypoplastic olfactory bulbs on the right and aplastic olfactory bulbs on the left. Eight patients with IA had bilaterally aplastic olfactory bulbs. The depth of the olfactory sulcus at the level of the PPTE was smaller in patients with IA than in control subjects. The depth of the olfactory sulcus was greater on the right than on the left, and there was no overlap. Among patients with IA, the depth of the olfactory sulcus differed significantly between those with and those without visible olfactory tracts.
CONCLUSION: The depth of the olfactory sulcus at the level of the PPTE reflects the presence of olfactory tracts. The presence or absence of the olfactory tract may therefore have some association with cortical growth of the olfactory sulcus region. The olfactory sulcus is deeper on the right than on the left, particularly in patients with IA. We speculate that olfaction may be processed predominantly in the right hemisphere.
Anosmia since birth or early childhood is an entity that can be divided into anosmias associated with chromosomal disorders (eg, Kallmann’s syndrome [1] and Klinefelter’s syndrome [2, 3]) or anosmias without evidence of other defects (isolated anosmia since birth or early childhood, termed “isolated anosmia” [IA] throughout this article). Although the underlying pathophysiology in Kallmann’s syndrome is relatively well investigated (4, 5), IA has received far less attention. However, the literature and our own clinical experience suggests that IA occurs more frequently than does Kallmann’s syndrome (6–9). For example, Leopold et al (8) reported that only two of 22 patients with “congenital anosmia” had either Kallmann’s syndrome or other endocrine defects. Similarly, six of seven patients investigated by Jafek et al (7) had no endocrine defects associated with anosmia. Potential causes may include genetic defects, infections, or trauma during late pregnancy or early childhood.
The present study was performed to investigate morphologic differences of frontobasal structures between patients with IA and control subjects. In addition, because volumetrics of the olfactory bulb seem to be of limited diagnostic value in individual patients (9–11), we also searched for a measure that would allow identification of IA more precisely than has been possible.
Methods
Patients
Twenty-one consecutive patients (19 female, two male; age range, 12–73 years; mean age, 29 years) were evaluated. They either were referred to the Department of Otorhinolaryngology at the University of Dresden by other university departments (eg, pediatrics and neurology) or presented themselves to the smell dysfunction clinic of the Department of Otorhinolaryngology. All subjects were in good health, with no signs or symptoms, except for anosmia. On careful questioning, none of these patients could remember any odorous sensations apart from intranasal sensations mediated by the trigeminal nerves (12–14). Specific questions about the distribution of axillary and pubic hair, fertility, and adult body development were asked to exclude hormonal problems that often are associated with congenital olfactory loss (see Introduction). Because none of the subjects had any indication of hormonal deficits, the chromosomal test for Kallmann’s syndrome was omitted. None of them had any kind of cranial malformation as verified by T1- and T2-weighted images of the whole brain.
Anosmia versus normosmia was ascertained psychophysically by using “Sniffin Sticks” (15, 16). The criterion for “functional anosmia” with the Sniffin Sticks was established in a study of 70 anosmic subjects in whom the diagnosis was made by means of detailed history, self-ratings of olfactory sensitivity, and olfactory evoked potentials (16). None of the anosmic subjects investigated in this study scored higher than 15. The test-retest reliability of the Sniffin Sticks test battery is 0.72 (15), which compares well with other tests of olfactory function (17). In addition to psychophysical testing, all anosmic patients also underwent electrophysiologic testing with chemosensory evoked potentials (18); none of the IA patients had responses to olfactory stimuli, whereas all of them responded to the trigeminal stimulant CO2.
We could perform MR imaging in 16 (14 female, two male patients; age range, 12–51 years; mean age, 27 years) of the 21 patients with IA. Their results were compared with those from eight control subjects who had no evidence of any disorder (four female, four male subjects; age range, 21–27 years; mean age, 24 years).
This study was approved by the institutional review board, and informed consent was obtained from all participants.
MR Imaging
MR imaging was performed by using a 1.5-T system (Magnetom Vision; Siemens Medical Systems, Erlangen, Germany) with a gradient strength of up to 15 mT/m; a standard quadrature head coil was used. After the scout images, a coronal T1-weighted spin-echo (SE) sequence was performed with the following parameters: 600/20 (TR/TE); section thickness, 3 mm (contiguous sections); matrix, 512; and field of view, 220 × 200 mm, resulting in a pixel resolution of 0.39 × 0.43 mm. Sections were angulated perpendicular to the anterior base of skull or cribriform plate. The observed region started at the frontal paranasal sinus and ended dorsally behind the crura cerebri. This was followed by a T1-weighted 3D magnetization-prepared rapid-gradient echo (MP-RAGE) imaging sequence with the following parameters: 9.7/4 (TR/TE); flip angle, 12°; and isotropic voxels, 1.0 × 1.0 × 1.0 mm. The entire brain was imaged with this sequence. Multiplanar reformation was conducted in three orthogonal planes.
Imaging Data Analyses
Acquired data sets were transferred to a workstation that had multiple measurement features (ID.Station, Read 3.6). Since standards of measurement for the olfactory bulbs, tracts, and sulci do not exist, acquired data were analyzed as follows: we measured volumes of the olfactory bulbs in patients with visible olfactory bulbs and in control subjects. The end of the olfactory bulb or beginning of the olfactory tract was seen clearly as a change in diameter, and boundaries of the olfactory bulbs were determined by the surrounding CSF and the anterior cribriform plate. The results of the bulb measurements in the control subjects were defined as normal; any bulb with a measured volume smaller than half the volume of the average of the normal bulbs was defined as hypoplastic. If no bulb was visible, we recorded aplasia of the bulb. Volumes of the olfactory tracts were not measured. The maximum length and depth of the olfactory sulcus on both sides were measured in all study subjects.
Because voxel size on the T1-weighted SE images was 0.5031 mm3 compared with 1 mm3 on the MP-RAGE images, and because the structures of interest ran perpendicular to the 2D images, we used the T1-weighted images to measure the volume of the olfactory bulbs and maximum depth of the olfactory sulci. Length of the olfactory sulci was measured along the axial reformation of the 3D data sets to reduce influences of partial-volume effects at the end of the olfactory sulci on the 2D images. Quantitative and qualitative analyses were performed by three readers (N.D.A., V.H., T.H.), and consensus was reached on the basis of manual segmentation.
To render the data less biased in terms of head size, relative lengths and depths of the olfactory sulci were computed as percentages of head size. For this, the maximum inner coronal diameter of the anterior base of the skull was measured. In addition, measurements of the depth of the olfactory sulci were performed in all subjects in a selected section of the acquired coronal T1-weighted SE images. To confirm identical section positions for this measurement, the most posterior coronal section through the eyeballs was used; on at least one side, a partial volume cut of the eyeballs had to be visible. This section was named the “plane of the posterior tangent through the eyeballs” (PPTE) (Fig 1).

Transverse reformation of a 3D data set of an MP-RAGE image. The eyeballs and olfactory bulbs are visible. The coronal plane indicates position of the plane of the PPTE. Note that in normosmic subjects with normal olfactory bulb, this plane cuts through the olfactory bulb.
By using the 3D data sets, qualitative analyses of asymmetries and volume loss were performed by three readers in consensus (N.D.A., V.H., T.H.) with regard to frontal and temporal lobes, prepiriform cortex, periamygdaloid area, and piriform lobe.
Statistical Analyses
Data were analyzed with SPSS 9.0 for Windows. For comparisons between groups of patients, multivariate analysis of variance was employed (analyses of variance, repeated-measures design; within-subject factor “side,” between-subject factor “group”). The df were adjusted according to Greenhouse-Geisser method. The α level was set at .05.
Results
Presence of Olfactory Bulb and Tract
All control subjects had symmetric olfactory bulbs (mean volume, 125.2 mm3 ± 17.2) and olfactory tracts (Fig 2). Olfactory tracts were visible from the end of the olfactory bulb to the anterior perforated substance. The olfactory bulb was absent bilaterally in eight patients with IA (50%); one of those patients had bilateral olfactory tracts (Fig 3), whereas the remaining seven did not show olfactory tracts (Fig 4). Five Patients with IA (31%) exhibited bilateral hypoplastic olfactory bulbs (mean volume, 18.5 mm3 ± 2.8), whereas all of them had olfactory tracts. In three patients (19%), we found unilateral hypoplasia of olfactory bulbs on the right (mean volume, 22.1 mm3 ± 6.0) and aplasia of the olfactory bulbs on the left. Correspondingly, in these patients the olfactory tracts were detectable only on the right and were absent on the left (Fig 5A and B).

Coronal T1-weighted SE image in the PPTE in a healthy subject. Note visualization of the olfactory bulbs and normal development of the olfactory sulci.
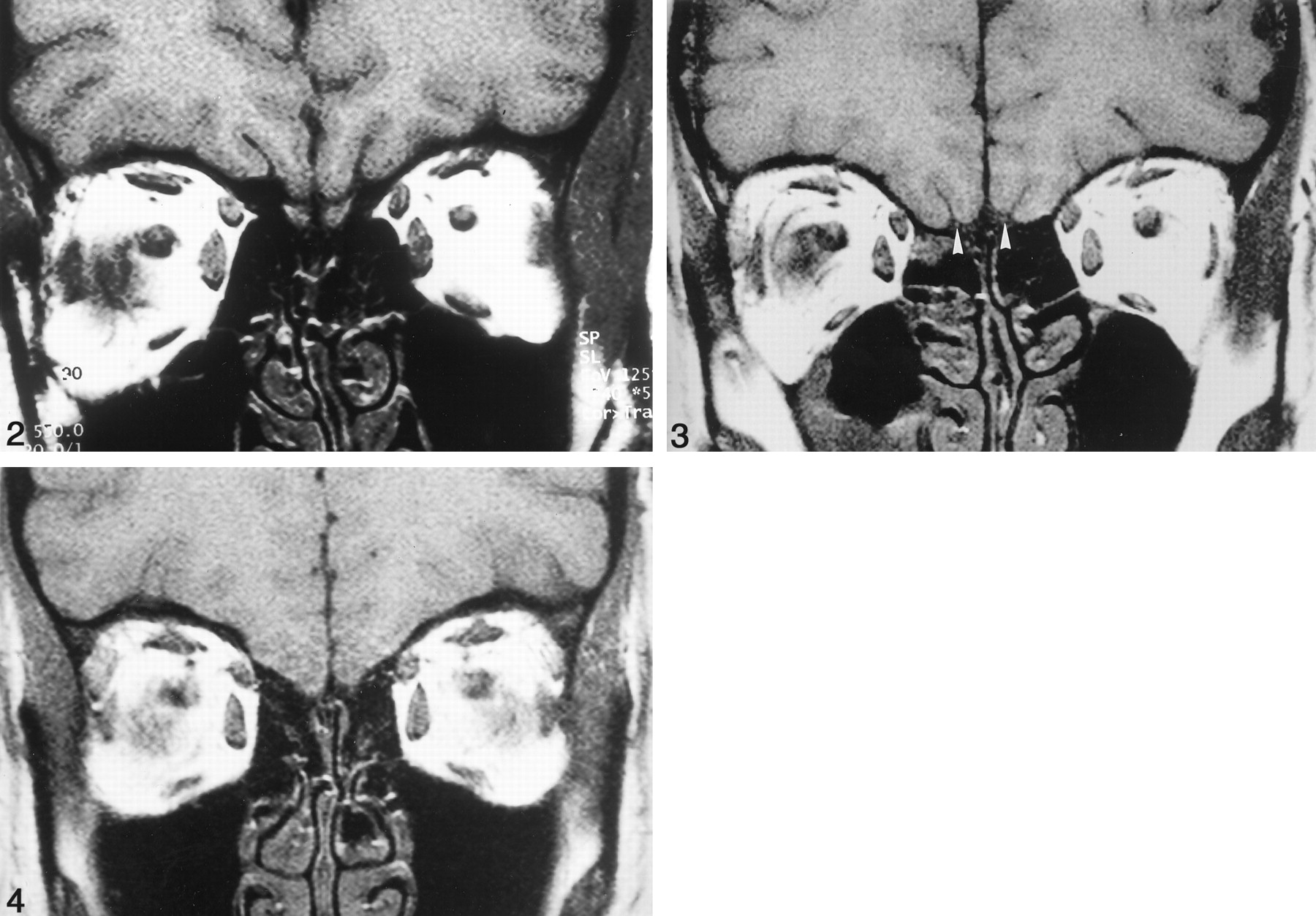
Coronal T1-weighted SE image in the PPTE in a patient with bilateral aplasia of the olfactory bulb, visible olfactory tracts (arrowheads), and slightly flattened olfactory sulci. Note partial volume effect of the right eyeball.

Coronal T1-weighted SE image in the PPTE in a patient with absent olfactory tracts and sulci. Note partial volume effects of both eyeballs.

Patient with olfactory tract present only on the right.
A, Coronal T1-weighted SE dorsal image in the PPTE. Olfactory tract is visible on the right (arrowhead). There is accordingly different development of the olfactory sulcus, as shown in B.
B, Transverse reformation of a 3D data set of an MP-RAGE image in the same patient. Olfactory tract on the left is absent; olfactory tract on the right is visible. Accordingly the olfactory sulcus is shorter (arrowhead) on the left where the olfactory tract is not detectable. Note exact transverse reformation as identified by symmetrical display of periorbital fat and the middle cerebral arteries.
The volume of hypoplastic olfactory bulbs varied from 14.7 to 26.8 mm3, with a mean of 19.4 mm3 ± 4.1. In patients with unilateral olfactory bulb aplasia, only the volume of the hypoplastic olfactory bulbs was included in the statistical analysis. In patients with visible olfactory bulbs (ie, no aplasia of the olfactory bulb), measurements revealed a mean volume of 20.2 mm3 ± 4.5 (maximum, 28.8 mm3; minimum, 14.7 mm3) for the right and 18.1 mm3 ± 2.1 (maximum, 20.1 mm3; minimum, 15.3 mm3) for the left.
Length and Depth of Olfactory Sulcus
Descriptive statistics of the olfactory sulcus measures are given in Table 1. Data from IA patients with measurable olfactory sulci were compared with those from control subjects. For the depth of the olfactory sulci in the PPTE, we found significant effects for both factors “side” (F1,20 = 7.9, P = .01) and “group” (F1,20 = 5.6, P = .03), with the right olfactory sulci being deeper than the left olfactory sulci and control subjects having deeper olfactory sulci than IA patients. In addition, there was a significant interaction between factors “side” and “group” (F1,20 = 6.8, P = .02), such that the difference in olfactory sulcus depth between the left and the right sides was much more pronounced in IA patients than in control subjects (Fig 6). In fact, most patients with IA who had detectable olfactory sulci in the PPTE had a deeper olfactory sulcus on the right. In control subjects, the right olfactory sulcus in the PPTE was found to be deeper than the left in three subjects (Table 2). Similar results were found for measures of olfactory sulcus depth in the PPTE relative to head circumference (factor “group”: F1,20 = 7.5, P = .01; interaction “side” × “group”: F1,20 = 9.0, P = .01).

Olfactory sulcus (OS) depth in PPTE in patients with IA compared with that in healthy controls (means, SEM). Only olfactory sulcus depths greater than 0 were taken into consideration. The olfactory sulcus was significantly deeper on the right than on the left.
Descriptive statistics of olfactory sulcus measures, both as absolute measures and as measures in relation to the maximum inner coronal diameter of the anterior skull
Depth of left and right olfactory sulci in control subjects and IA patients with a visible olfactory sulcus in the PPTE
Except for significantly larger maximum depth of the olfactory sulcus on the right compared with the left (relative measures, factor “side”: F1,22 = 6.6, P = .02), significant differences were not observed for either measure of the olfactory sulci (maximum olfactory sulcus length, factor “side”: F1,22 = 1.25, P = .28, factor “group”: F1,22 = 0.05, P = .82; relative olfactory sulcus length, factor “side”: F1,22 = 1.20, P = .29, factor “group”: F1,22 = 0.01, P = .92; maximum olfactory sulcus depth, factor “side”: F1,22 = 2.65, P = .12, factor “group”: F1,22 = 0.55, P = .47; relative maximum olfactory sulcus depth, factor “group”: F1,22 = 0.33, P = .57).
Relationship of Depth of Olfactory Sulci in the PPTE to Visibility of Olfactory Tracts
When analyzing the depth of the olfactory sulci in relation to the visibility of the olfactory tracts, we divided the cases into two groups: visible olfactory (VOT) tracts ([VOT] (n = 21) and nonvisible olfactory tracts ([NOT] n = 11) (Table 3). No overlap existed between the two groups with regard to the depth of the olfactory sulci in the PPTE (Fig 7). The depth of the olfactory sulcus in the PPTE in the VOT group ranged from 5 to 12 mm (mean, 7.9 mm ± 1.6), and in the NOT group, from 0 to 4 mm (mean, 1.6 mm ± 1.4).
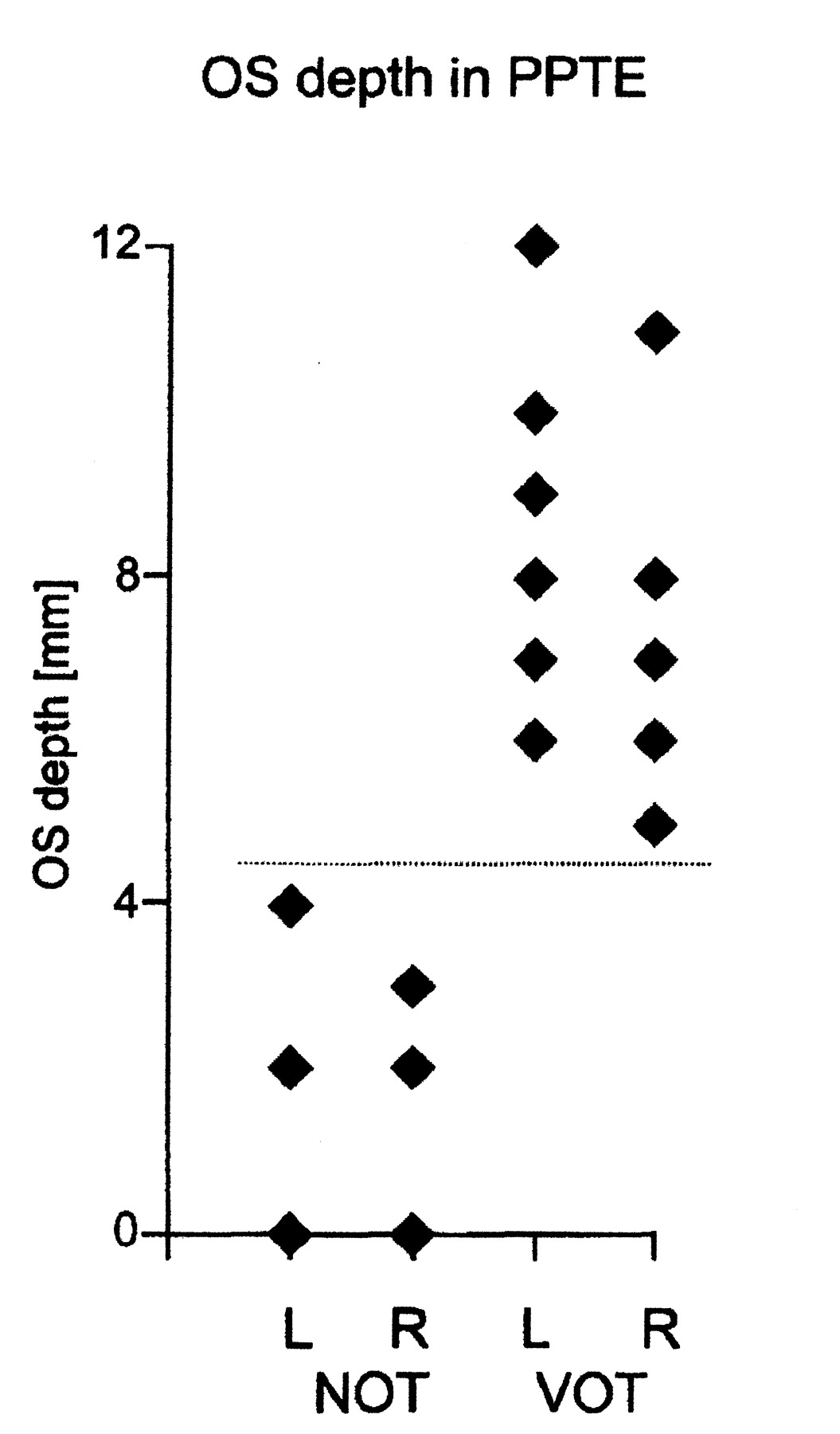
Olfactory sulcus (OS) depth in the PPTE in IA patients with visible olfactory tracts (VOT) and in those with nonvisible olfactory tracts (NOT). There is no overlap between the two groups in terms of olfactory sulcus depth.
Olfactory sulcus depth in the PPTE in patients with IA by presence of olfactory bulb and olfactory tract
Qualitative Analyses
By consensus, qualitative examination of the frontal and temporal lobes, prepiriform cortex, periamygdaloid area, and piriform lobe revealed no differences between patients with IA and normosmic control subjects. Specifically, patients with IA showed neither asymmetries nor volume loss.
Discussion
The present investigation yielded three major findings: 1) the olfactory sulcus in the PPTE is significantly less deep in patients with IA; 2) the olfactory sulcus is significantly deeper on the right than on the left, and this difference is more pronounced in patients with IA than in normosmic control subjects; and 3) in our sample, no overlap existed in the depth of the olfactory sulcus in the PPTE in patients with IA with olfactory tracts (larger olfactory sulcus) and those without olfactory tracts (smaller olfactory sulcus).
To our knowledge, detailed measurements of the olfactory sulci in patients with IA have not been published previously. In congenital anosmia, however, the length and depth of the olfactory sulcus have been reported to be aplastic, hypoplastic, or normal (9, 18–22). In the present study, measurements of the depth of the olfactory sulcus differed consistently between patients with IA and control subjects only when performed on a coronal image perpendicular to the frontal skull base posterior to the eyeballs (PPTE). Measurements did not differ significantly between those in control subjects and those in patients with IA when maximum values of the olfactory sulcus for either length or depth were taken into consideration. Accordingly, aplasia or hypoplasia of the olfactory sulcus now can be quantified.
In the PPTE, all patients with visible olfactory tracts had an olfactory sulcus depth >4 mm, whereas in patients with no visible olfactory tract, the olfactory sulcus depth was ≤ 4 mm. This compares with findings by Yousem et al (9) for Kallmann’s syndrome; they reported that “in 18 instances (out of 50 possible), a bulb was absent but an olfactory sulcus was present, and in no instance was an olfactory sulcus absent but an olfactory bulb present.”
On a diagnostic level, these findings indicate that measurement of the depth of the olfactory sulcus in the PPTE is a good indicator of the presence of olfactory tracts, which frequently are difficult to detect with standard imaging techniques. In addition, in daily routine imaging, the olfactory sulci are clearly visualized in axial and coronal planes with standard techniques. Because the olfactory bulbs and tracts are not clearly depicted with standard imaging techniques, detection of morphologic changes of the olfactory sulci can indicate an underlying IA with hypoplasia or aplasia of the olfactory bulbs.
On a developmental level, these findings may indicate that the formation of the olfactory sulcus, at least to some degree, depends on the presence of an olfactory tract (23). In addition, it is interesting to note that lateral aplasia of olfactory bulb and tract also was reflected in lateral depths of the olfactory sulcus. We speculate that this could reflect the CNS information processing of the olfactory system, which is thought to be more ipsilateral to the stimulated nostril, at least to a certain degree (24).
Our findings in IA compare with those reported for Kallmann’s syndrome. In both groups of patients, three different stages have been reported: 1) hypoplasia of the olfactory bulbs with olfactory tracts present, 2) aplasia of the olfactory bulbs with olfactory tracts present, and 3) aplasia of both olfactory bulbs and olfactory tracts (20, 21, 25). It remains unclear whether these similarities in phenotype are based on a similar defect present in both groups. In fact, although numerous studies have investigated genetic defects in Kallmann’s syndrome (4), to our knowledge, no such studies exist for IA. This is despite the impression that IA appears to occur more frequently than does Kallmann’s syndrome (7–9).
Similar to what has been described for Kallmann’s syndrome (26, 27), our qualitative analysis of the frontal and temporal lobes, prepiriform cortex, periamygdaloid area, and piriform lobe did not reveal major differences between patients with IA and control subjects. In Kallmann’s syndrome, however, the same authors also have described mild to moderate volume loss in temporal and frontal lobes (9). Thus, in Kallmann’s syndrome, changes in secondary or tertiary cortical centers involved in the processing of olfactory information appear to be less pronounced than are seen for areas that may be more directly involved in the higher-order processing of odorous sensations, such as the olfactory sulcus.
The differences between the depths of the left and right olfactory sulci in the PPTE were an interesting and, to some degree, unexpected finding of this study. The structural difference between the left and right hemisphere bears similarities to work in experimental animals, in which olfactory bulb volume has been shown to be larger on the right compared with the left (28). In contrast, in humans little support exists for such lateral differences in structures involved in the processing of olfactory information. Yousem et al (10) had no evidence for lateral differences at the level of the olfactory bulb or the temporal lobe, which is exactly what we observed in the present study. Despite these negative findings for the olfactory bulb and the temporal lobe, and although the lateral differences in olfactory sulcus depth were most pronounced in patients with no olfactory input, the left-right differences between depths of the olfactory sulci may relate to the higher-order processing of olfactory information. Specifically, accumulating evidence from psychophysical, clinical, electrophysiologic, and imaging studies show that, in general, the right hemisphere is more important to the perception of smells than is the left hemisphere (29–34). Further, the present data appear to indicate a relation between presence of an olfactory tract and depth of the olfactory sulcus in the PPTE. The presence or absence of the olfactory tract, therefore, may have some association with cortical growth of the olfactory sulcus region.
These speculations imply, at least in part, that the presence or absence of sensory input leads to structural changes at the level of the CNS. Such morphologic consequences of missing sensory input have been shown for vision (35), hearing (36, 37), and the somatosensory systems (38) with sensory deprivation during critical periods of embryologic development. In the olfactory system, for example, olfactory deprivation has been shown repeatedly to result in reduced volume of the olfactory bulb (39, 40). No such study exists for the olfactory sulcus in humans, however. In addition, to our knowledge, no studies exist on the embryologic development of the human olfactory sulcus.
Conclusion
The present study indicates that the depth of the olfactory sulcus in the PPTE provides additional information about the presence of olfactory tracts. In turn, this also may imply that the presence of olfactory tracts promotes cortical growth in the area of the olfactory sulcus despite the lack of neuronal input from a functional olfactory bulb. Finally, the right-sided lateralization of olfactory sulcus depth may indicate at a morphologic level that information in the olfactory system is predominantly processed in the right hemisphere.
Acknowledgments
We would like to thank Andrew Livermore, Charles Sturt University, Bathurst, Australia, for his thoughtful comments on this manuscript.
References
- Received March 14, 2001.
- Accepted after revision August 14, 2001.
- American Society of Neuroradiology
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Acquired olfactory loss alters functional connectivity and morphology
- Bilateral transient olfactory bulb edema during COVID-19-related anosmia
- Structural changes in secondary, but not primary, sensory cortex in individuals with congenital olfactory sensory loss
- Brain Changes in Kallmann Syndrome
- The Depth of the Olfactory Sulcus Is an Indicator of Congenital Anosmia
- Association of Olfactory Bulb Volume and Olfactory Sulcus Depth with Olfactory Function in Patients with Parkinson Disease