
Abstract
BACKGROUND AND PURPOSE: Bilateral vertebrobasilar junction agenesis is an exceptional anatomic variation. This article explores the angiographic characteristics of this variant and its embryologic mechanisms.
MATERIALS AND METHODS: Two observations of bilateral agenesis of the vertebrobasilar junction are reported. A case of atheromatous disease of the vertebrobasilar junction is shown to highlight characteristics distinguishing such a lesion from the reported variant.
RESULTS: In the 2 reported cases, the distal segment of both vertebral arteries (VAs) and the proximal portion of the basilar artery (BA) were absent. In addition, distal connections of the BA with the posterior cerebral arteries (PCA) were also lacking. As a consequence, the remaining portion of the BA was isolated from its usual sources of blood supply, which was provided by a persistent carotid-basilar anastomosis.
CONCLUSION: The developmental mechanism underlying bilateral agenesis of the vertebrobasilar junction likely involves the anterior radicular artery of C1. This branch of the proatlantal artery normally becomes the adult distal VA and the proximal BA. The lack of cranial connection of the BA with the PCA may be secondary to the proximal vertebrobasilar agenesis and the resulting paucity of antegrade flow within the BA. Alternatively, the absence of both the proximal and distal connections of the BA could be the result of a similar, yet unknown, developmental mechanism. From a clinical standpoint, this vascular anomaly was discovered incidentally in our 2 patients, a finding consistent with the assumed congenital nature of the variant.
Absence or hypoplasia of the terminal portion of 1 vertebral artery (VA) is a commonly observed anatomic variant. In such instance, the VA either shows a very small connection to an otherwise normal basilar artery (BA) or continues its course as the posterior inferior cerebellar artery (PICA). In both cases, the BA is normal and receives most or all of its blood supply from the contralateral VA. Absence of the terminal portion of both VAs is, on the other hand, exceptional. We report 2 angiographic observations of bilateral absence of the distal VA. Remarkably, in both cases, the proximal portion of the BA was missing as well, whereas the remaining segment of the BA was only connected to the anterior circulation through a persistent embryonic vessel. We believe these cases illustrate a congenital anatomic variation that can be explained by the developmental history of the vertebral and basilar arteries. A third case of vertebrobasilar junction disease that presents similarities with this anatomic variation is also reported, and the distinguishing features are discussed.
Case Illustrations
Case 1
A 69-year-old woman was investigated for a right pulsatile tinnitus. Her history was remarkable for arterial hypertension, migraine headaches, and premature birth (30-week gestation). MR imaging and angiography revealed a left persistent trigeminal artery (PTA), possibly associated with an aneurysm. Digital subtraction angiography (DSA) showed no aneurysm but documented several anatomic variations. Both posterior cerebral arteries (PCAs) originated from the distal internal carotid artery (ICA), without detectable connection with the BA. There was no evidence of connection between the VA and the BA on either side. The right VA terminated as a branch of the occipital artery, whereas the left VA crossed the midline to continue as the right PICA. The left PICA was provided by the left anterior inferior cerebellar artery (AICA) through a common stem (AICA-PICA trunk). Because of the absence of connection with both PCA and both VA, the BA was isolated from its usual sources of blood supply and was vascularized by the left PTA only (Fig 1A, -B). Finally, the anterior spinal artery originated from the right inferior thyroid artery (Fig 1C).

A 69-year-old woman with pulsatile tinnitus. A, DSA, left common carotid injection, lateral view, showing a PTA opacifying the BA. Note the common origin of the left PCA and left AchoA from the distal ICA. B, DSA, left common carotid injection, right anterior oblique view, showing the basilar artery fed by the PTA (arrowhead) and the bilateral absence of connection between the basilar and posterior cerebral arteries. The BA ends into the superior cerebellar arteries distally and into prominent AICA proximally. The left AICA also supplies the left PICA territory. The right PICA territory is fed by the left VA (not shown). There is no visible connection between the VAs and the BA. C, DSA, right subclavian injection, showing opacification of the anterior spinal artery (arrowheads) via the right inferior thyroid artery.
Case 2
A 65-year-old man followed for Wegener granulomatosis and right-ear deafness underwent MR imaging and angiography, which revealed a possible right ICA termination aneurysm. DSA showed no aneurysm but documented several vascular variations. On both sides, the VA continued as the ipsilateral PICA without connection to the BA. A prominent right anterior choroidal artery (AchoA) was supplying the territory of the right PCA, without connection to the BA (Fig 2A). The right posterior communicating artery (PcomA), separate from the PCA, was supplying the BA, which showed retrograde opacification down to the level of origin of both AICAs (Fig 2B). Again, there was no connection between the right PcomA and the right PCA itself. The left PCA originated from the distal left ICA without detectable connection to the BA. Because of the absence of connection with both PCAs and both VAs, the BA was isolated from its usual sources of blood supply and was fed through the right PcomA only.
A 65-year-old man with Wegener granulomatosis. A, DSA, right common carotid injection, lateral view, showing a prominent AchoA supplying the right PCA territory (arrowhead). Note a separate right PcomA feeding the BA (arrow). B, DSA, right common carotid injection, transfascial view, showing the BA fed by the right PcomA. The BA ends into the superior cerebellar arteries (SCAs) distally and into the AICA arteries proximally. Both VAs end as ipsilateral PICAs without connection to the BA (not shown).
Case 3
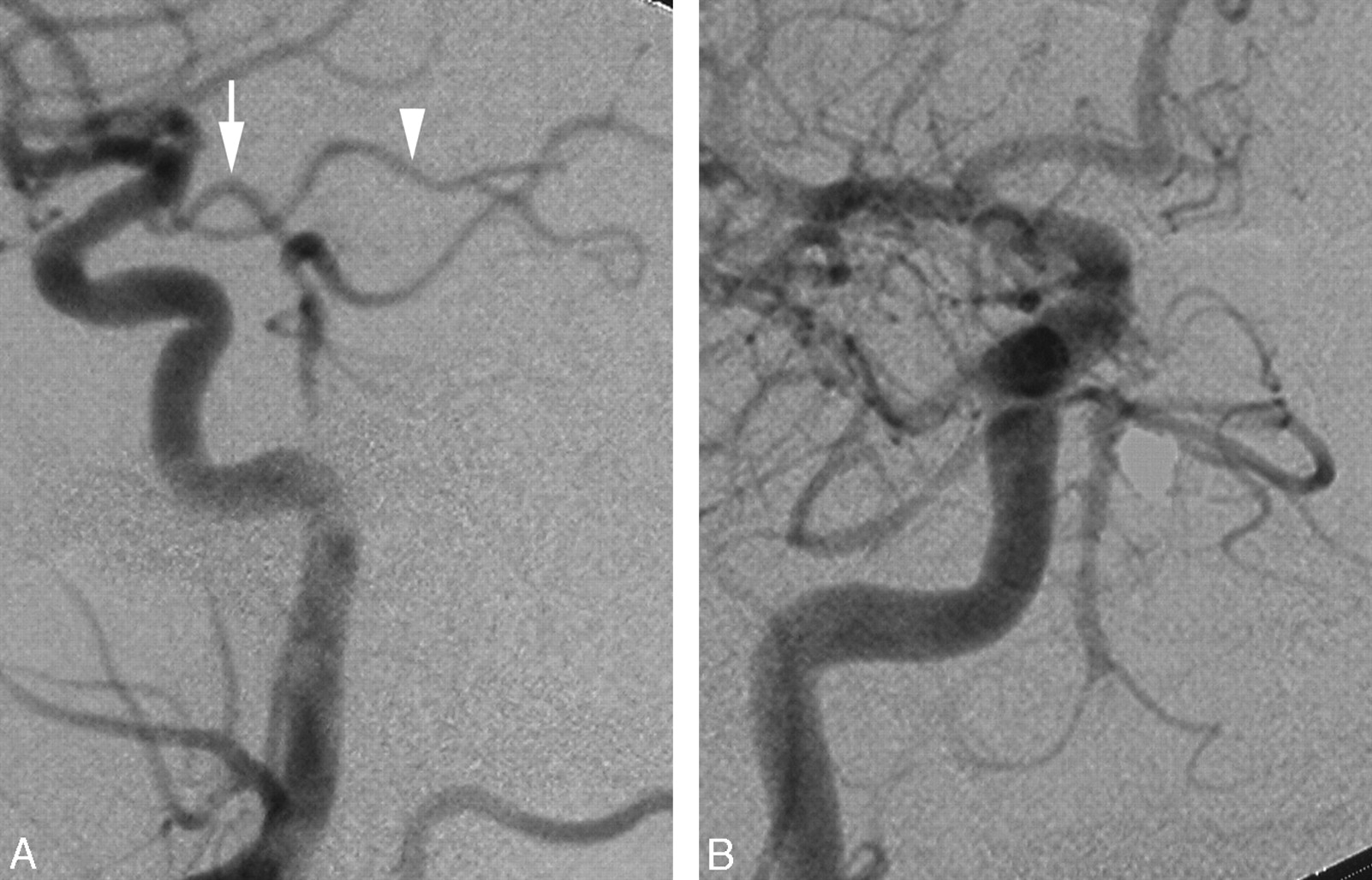
A 51-year old man with a history of coronary artery disease, arterial hypertension, hyperlipidemia, and smoking presented with a 1-year history of episodic dizziness associated with a throbbing sensation in both ears, elicited by head rotation to the right or left. Initial investigation for benign paroxysmal positional vertigo remained inconclusive. MR imaging and angiography offered only poor visualization of the BA and VA. DSA showed that the extracranial portion of both VAs was of small caliber but otherwise unremarkable. The right VA was not connected to the BA but continued as the vermian branch of the right PICA (not shown), whereas the hemispheric branch took origin from the right AICA. The V4 segment of the left VA, though small and irregular, was seen joining the proximal BA, which showed antegrade opacification up to the level of origin of both AICAs (Fig 3A). The vermian and hemispheric branches of the left PICA were both arising from the left V4 segment. The left ICA contributed to the BA via a large PcomA, which itself continued as the left PCA (Fig 3B, -C).

A 51-year-old man with intermittent dizziness. A, DSA, left VA injection, anteroposterior view, showing disease in the distal VA and proximal BA (arrowhead), including faint opacification of the right AICA (arrow). B, DSA, left common carotid artery injection, lateral view, showing a large PcomA supplying the BA. Note that the PcomA then continues as the left PCA (arrowhead). The hemispheric branch of the right PICA territory is fed through a connection with the BA. C, DSA, left common carotid injection, transfascial view, showing a large left PcomA supplying the BA and continuing its course as the left PCA. Note the opacification of the right AICA (arrow), which confirms the existence of a connection between the left VA and the distal BA.
Discussion
Developmental Anatomy of the Vertebrobasilar Junction
The VA develops as a series of longitudinal anastomoses established between the sixth intersegmental artery (ie, the future subclavian artery) and the proatlantal artery of Padget (ProA).1,2 Because of this peculiar mode of formation, the VA has been described as a “hemodynamic solution” rather than a true vessel.3 The VA in its adult form incorporates a portion of the ProA. Initially, the ProA has a typical segmental configuration, with a dorsospinal division that sends a radicular branch along the first cervical nerve root (C1). At the adult stage, this radicular artery of C1 becomes the terminal segment of the VA.4 It divides into anterior and posterior radicular branches just before entering its intradural course. The anterior radicular branch turns, therefore, into the intradural component of the VA (V4 segment) plus a short extradural segment (distal V3 segment). Distally, it divides into ascending and descending rami that fuse along the midline with their contralateral counterparts to form the BA and the anterior spinal axis, respectively. The descending ramus of the posterior radicular branch of C1 becomes the ipsilateral cranial origin of the posterior spinal artery,5 whereas its ascending ramus establishes a small connection with the ipsilateral PICA. From a phylogenetic standpoint, the role of the arterial branch traveling along the first cervical nerve route can be traced back to snakes, in which a substantial contribution to the lower BA is provided at the C1 level, while the VA per se has not developed yet.6 In fact, Gillilan6 noted that “the artery accompanying the first cervical nerve is conspicuously large in all snakes studied.” This C1 contribution can be found in other reptiles as well, such as the alligator.7 Despite the profound modifications undergone by the embryonic pattern at the cervical level, the basic configuration can, therefore, still be read in terms of typical segmental spinal vascularization (Fig 4).

Schematic representation of the vertebrobasilar circulation. Posterior view of the VAs and BA. On the left side, the segments derived from the radicular branch of the proatlantal artery are shown in gray. Ant. Spinal a. indicates anterior spinal artery; Post. Spinal a., posterior spinal artery.
Bilateral Vertebrobasilar Agenesis
The anatomic variation presented here is characterized by a set of angiographic findings that include bilateral absence of connections between the BA and the PCA cranially and between the BA and the VA caudally. As a result, the BA is, in both reported cases, isolated from its usual sources of blood supply, in a manner reminiscent of its embryonic state before the connections with the PCA and the terminal VA are established. It is, therefore, not surprising to find that the blood supply to the BA is provided, in our patients, by a persistent embryonic vessel, (ie, a PTA in case 1 and a PcomA in case 2). The role played by persistent carotid-basilar anastomoses, in particular by PTAs, as sources of collateral supply in cases of congenital anomalies of the carotid and vertebral axes has been well described.8,9 It may be useful to mention here that the PcomA should be considered, in the fetus, as the most cranial of the carotid-basilar anastomoses rather than as the caudal division of the fetal ICA.10 In a superb study dealing with the development of the PCA in the rat, Moffat11 has shown that the territory of the PCA is first provided by the AchoA (not by the PcomA, as often believed) and is later transferred to the cephalic end of the ipsilateral longitudinal neural artery (ie, the future BA) through the formation of a connection between the AchoA/PCA and the posterior choroidal artery (PchoA), itself a branch of the PcomA (Fig 5A–D).

Developmental anatomy of the distal BA. A, The anterior circulation supplies the developing posterior circulation (ie, the parallel longitudinal neural arteries [LNA]) via the trigeminal and the posterior communicating arteries (PcomAs). The terminal branches of the carotid axis are the AchoA, which provides the territory of the PcomAs and the anterior cerebral artery (ACA) and its developing branch, the middle cerebral artery (MCA). The VA is not yet connected to the LNA. The PcomA has 2 small branches, the PchoA and the diencephalic artery (Dienceph. A). A indicates artery. B, The VA connects to the proximal end of the LNA/BA. The trigeminal artery involutes, and the PcomA becomes the last of the carotid basilar anastomoses. C, A connection is formed between the AchoA/PCA and the PchoA. D, The connection between the PcomA and the PCA is now well established. The AchoA is not supplying the PCA territory anymore. The latter can be supplied equally by the posterior circulation via the distal segment of the PcomA or by the anterior circulation via the proximal segment of the PcomA. This configuration of equal contribution can persist at the adult stage. Other adult configurations depend on the relative regression of the PcomA, as described in E and F. Note that the diencephalic branch is now called the posterior medial choroidal artery (PMchoA), whereas the original PchoA is now called the posterior lateral choroidal artery (PLchoA).11 Although both arteries are originally derived from the PcomA, they are considered as branches of the PCA at the adult stage. The so-called “P1 segment of the PCA” is in fact the distal portion of the PcomA. E, In this second adult configuration, the distal segment of the PcomA has regressed and the anterior contribution to the PCA becomes dominant. This configuration is often mislabeled as a “fetal origin of the PCA.” The true “fetal origin of the PCA,” where the transfer of the PCA territory from the AchoA to the PcomA has not occurred, is not illustrated here but would be similar to B. It is illustrated in Fig 2A. F, In this third adult configuration, the proximal segment of the PcomA has regressed and the posterior contribution to the PCA becomes dominant.
At the adult stage, the PCA territory is controlled by the BA or by the PcomA, depending on the relative degree of regression of the PcomA proximal and distal to the connection established with the AchoA/PCA (Fig 5E–F). This developmental pattern is consistent with modern observations made on human fetuses.12 It therefore appears that the segment of PCA located, in the adult configuration, between the PcomA and the BA, and classically named the P1 segment of the PCA, is in fact a portion of the PcomA located distal to the newly established connection with the AchoA/PCA. It also becomes clear that, at the adult stage, a dominant PcomA controlling the PCA territory should not be called a fetal origin of the PCA. This appellation should be reserved for the less common variant that sees the PCA territory provided by the AchoA. Our second case offers a “snapshot” of this early developmental stage, in which the right AchoA continues as the PCA, whereas a separate right PcomA connects to the BA as a typical carotid-basilar anastomosis. In this case, the 2 vessels can be clearly labeled because they are simultaneously present. The distal point of the origin of the other 3 PCAs seems compatible with this variant as well, (ie, an AchoA origin of the PCA), but definitive identification is, in these cases, hindered by the lack of a detectable PcomA. The absence of connection between the BA and the PCA in each instance is consistent with the hypothesis that the PCA territory has not been transferred from the AchoA to the longitudinal neural artery. Again, the presence or absence of a so-called P1 segment located between the PcomA and the BA is irrelevant because this segment, in fact, corresponds to the distal portion of the PcomA. The vascular segment important to this discussion, whose presence or absence decides the fate of the PCA territory, becomes, at the adult stage, a portion of the PCA located distal to its connection with the PcomA.
Another important observation made in both our cases is the absence of the proximal portion of the BA in addition to the bilateral lack of a distal VA. This finding comes, in fact, as a logical consequence of the developmental anatomy of the vertebrobasilar junction. As mentioned previously, the proximal BA results from the fusion of the ascending rami of the anterior radicular branches of C1, originating from the left and right ProAs. Bilateral agenesis of these anterior radicular branches, therefore, explains the absence of the proximal BA as well as the absence of both distal VAs (Fig 4). On the other hand, agenesis of a single anterior radicular branch would only result in the absence of 1 distal VA, the contralateral VA, and the BA being unremarkable, a variant that is in fact commonly observed.
The PICA normally arises from the portion of the VA that is missing in our 2 patients. The origin of the PICA in these cases documents the use of previously described collateral pathways. In the first patient, the left PICA comes from the BA as a common stem with the left AICA (AICA-PICA trunk), whereas the right PICA is the continuation of the left VA. In the second patient, each PICA is the cranial continuation of the ipsilateral VA. A proximal origin of the PICA from the VA can be understood either as a variation of the lateral spinal artery,13 or as a variation of the posterior spinal artery.14 In the latter instance, the PICA is a branch of the posterior spinal artery via the connection normally established between these 2 vessels by the ascending ramus of the posterior radicular branch of C1. This suggests that the cause of bilateral vertebrobasilar junction agenesis, as seen in our 2 patients, may be limited to the congenital absence of the anterior radicular branch of C1.
It is conceivable, though not proved, that the absence of a connection between the BA and the PCA comes, in our 2 cases, as a secondary consequence of the lack of proximal connection with the VA and the resulting paucity of antegrade flow within the BA. Alternatively, the simultaneous lack of connections between the BA and the PCA cranially and the BA and the VA caudally could be related to a common, yet unknown, mechanism interfering with the development of such vascular segments at a specific moment of the fetal life. A mechanism involving the secondary regression of previously existing vascular segments seems unlikely in view of the well-defined “segmental” pattern of the vascular anomaly and the persistence of an embryonic branch as source of collateral blood supply. These characteristics may be compared with reported cases of bilateral segmental regression of the carotid and vertebral arteries, in which collateral supply was provided by the development of vascular retia rather than by an embryonic artery.15,16 As emphasized by Mahadevan et al, 15 the development of retia as collateral pathways indicate regression of a previously existing vessel rather that an archaic variation.
Finally, in the first patient, the anterior spinal axis received a prominent contribution from the right inferior thyroid artery. This variant is rare.17 It may be seen here as a collateral pathway developed to palliate the absence of cranial contributors of the anterior spinal axis from the distal VA (ie, the descending ramus of the anterior radicular artery of C1).
Literature Review
A review of the literature revealed only 1 observation similar to the variant presented here. Hoh et al18 have recently described a case of “persistent axial nonfusion of the basilar artery.” In this case, as in our first patient, each VA continues as the ipsilateral PICA, and the BA, which is supplied by a PTA, seems absent below the level of the AICA. The authors attributed the vascular anomaly to “a failure of fusion of the distal basilar artery,” to a “midbasilar agenesis,” and to “the PICA termination of the VA.” We believe that their variant is similar to the one documented in our 2 patients and that it can be explained by the developmental mechanism we have described previously (ie, the agenesis of the radicular artery of C1 or of its anterior radicular branch).
Clinical Presentation and Implications
The 2 patients with vertebrobasilar agenesis described in this report were investigated for an unrelated reason (ie, a pulsatile tinnitus in the first patient and Wegener granulomatosis in the second). In each patient, MR angiography was suggestive of intracranial aneurysm, which led to obtaining a diagnostic angiogram. There was no sign or symptom that could be directly referred to the vascular anomaly in either patient. It can, therefore, be said of their anatomic variants that they were discovered incidentally. This is not the case of our third patient, whose clinical presentation was suggestive of vertebrobasilar insufficiency. The morphology of the vascular anomalies revealed by DSA in this patient was, moreover, pointing toward a pathologic process rather than a congenital variant. The principal feature that helps distinguish the 2 situations is the presence of a connection between the left VA and the BA. The small and irregular aspect of the distal left VA was consistent with an atheromatous lesion, whereas the proximal portion of the BA, though abnormal in appearance, was present. The fact that the left PcomA, in addition to providing collateral supply to the BA, was controlling the ipsilateral PCA territory is of particular interest in regard to the pattern of development discussed earlier. It indicates that the fetal transfer of the PCA territory from the AchoA to the PcomA did occur normally in this patient, as opposed to the previous 2 patients.
Conclusions
In summary, we present 2 cases of selective bilateral agenesis of the vertebrobasilar junction. This anatomic variation is characterized by the absence of the distal portion of both VAs as well as the absence of the proximal portion of the BA. The lack of cranial connection of the BA with the PCA further isolates this vessel from its normal sources of blood supply. As a consequence, the BA is controlled by a persistent carotid-basilar anastomosis such as a PTA or a PcomA. The mechanism underlying this variant may be as limited as the agenesis of the anterior radicular artery of C1, a branch of the ProA that normally becomes the distal segment of the VA and the proximal BA at the adult stage. The lack of cranial connection of the BA with the PCA could be understood as a remote consequence of the proximal vertebrobasilar agenesis and the resulting paucity of flow within the BA. Alternatively, it is possible that a similar, yet unknown mechanism is responsible for the absence of both the proximal and distal connections of the BA. From a clinical standpoint, this vascular anomaly was discovered incidentally in our 2 patients, a finding consistent with the supposed congenital nature of the anatomic variant.
References
- Received November 21, 2006.
- Accepted after revision April 26, 2007.
- Copyright © American Society of Neuroradiology





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}