
Abstract
BACKGROUND AND PURPOSE: Nowadays the mechanism of homosexuality is little known. Few studies have been carried out to explore the brain functional changes of homosexual men during sexual arousal. We used functional MR imaging (fMRI) to determine whether the patterns of brain activation in homosexual and heterosexual men differed during visually evoked sexual arousal.
MATERIALS AND METHODS: To all the subjects (10 homosexual and 10 heterosexual), real-time visual stimulation was provided by 3-minute exposure to 3 types of erotic film: heterosexual couples (F-M), male homosexual couples (M-M), and female homosexual couples (F-F) engaged in sexual activity, during which time fMRI was used to determine the patterns of brain activation. Self-reports of level of sexual arousal were collected immediately afterward.
RESULTS: Statistical parametric mapping showed that viewing erotic film excerpts that induced sexual arousal was associated, in both groups, with activation of the middle prefrontal gyrus, bilateral temporal lobe and postcentral gyrus, thalamus, insula, vermis, left precuneus, occipital cortex, parietal cortex, and cerebellum. In homosexual men, the left angular gyrus, left caudate nucleus, and right pallidum were activated; in contrast, heterosexual men showed no activation in these regions. However, heterosexual men showed activation in the bilateral lingual gyrus, right hippocampus, and right parahippocampal gyrus, areas not activated in homosexual men. In both groups, region-of-interest analysis revealed no correlation between the magnitude of amygdala or thalamus activation and the reported level of sexual arousal.
CONCLUSION: Our findings indicate that different neural circuits are active during sexual arousal in homosexual and heterosexual men and may contribute to a better understanding of the neural basis of male sexual orientation.
Sexual orientation refers to sexual attraction toward persons of the opposite sex or the same sex and consists of selection and preference mechanisms sensitive to sex, with motivational approach behaviors toward the preferred individual. In the past several decades, these ideas have guided investigators to search for the possible origins of human sexual orientation in androgens and their target neural substrates. Within this framework, one hypothesis suggests that heterosexual preference in men is due to typical prenatal exposure to androgens, whereas homosexuality in men is due to underexposure to prenatal androgens.1 Recently, research in this area has focused on markers such as the ratio of the second-to-fourth finger lengths.2 However, the maternal immunity hypothesis is certainly the most revolutionary neurodevelopmental model of human sexual orientation. It rests on 1 very reliable finding, the fraternal birth order effect—that is, homosexual men have a greater number of older brothers than heterosexual men.3
We still know very little about differences in the neural circuits between people with same-sex attraction and those with opposite-sex attraction. Simon LeVay's (1991) autopsy study showed that the third interstitial nucleus of the anterior hypothalamus (INAH-3) is smaller in homosexual men than in presumed heterosexual men and indistinguishable from that in presumed heterosexual women.4 However, another study found a nonsignificant trend for a female-typical INAH-3 among homosexual men.5 One recent positron-emission tomography (PET) study demonstrated a stronger hypothalamic response to serotonergic challenge in heterosexual than in homosexual men6; and neuroimaging studies comparing heterosexual men and women while viewing preferred sexual imagery showed significantly greater hypothalamic activation in heterosexual men.7 These findings could be taken to suggest that there is a functionally distinct hypothalamic substrate for sexual attraction toward women.
The present functional MR imaging (fMRI) study was undertaken to compare the neuroanatomic correlates of visually evoked sexual arousal in healthy homosexual and heterosexual men; the study was largely exploratory.
Materials and Methods
Subjects
Subjects were enrolled through advertisements on campus and in a homosexual club asking for participation in a study on central nervous processing of sexual stimuli. Twenty healthy men took part in the study, 10 homosexual (mean age, 26.50 ± 5.10 years) and 10 heterosexual (mean age, 27.90 ± 3.03 years). The sexual orientation of all subjects was assessed by using self-reported measures of sexual feeling (ie, sexual attraction and sexual fantasies). In a structured interview, we verified that participants had no history of substance abuse, sexual dysfunction, sex identity disorder, paraphilia, or sexual offenses. All subjects gave informed consent to participate, and the study was approved by the Zhejiang University Human Investigations Committee. All subjects were told in detail about the aims of the study and what was expected of them in the experiment.
Stimuli and Procedures
Real-time visual stimulation was provided by alternately combining 3 types of erotic film: heterosexual couples (F-M), male homosexual couples (M-M), and female homosexual couples (F-F) engaged in explicit sexual activity and a visual fixation cross. Each type of erotic film was montaged with attractive short films.
Stimuli were presented via presentation software (Neurobehavioral Systems, Albany, Calif). No stimulus was repeated during the experiments. The sexual stimulation paradigm consisted of 3 alternating periods, with a visual fixation cross-presented for 3.0 minutes followed by each erotic film presented in pseudorandom order for 3.0 minutes. A mirror placed in front of the subject's eyes enabled him to see the stimuli presented on a screen located 2 m from his head.
Ratings of sexual attractiveness were assessed immediately after each scan. Homosexual and heterosexual men judged the sexual effect of the stimuli by means of a Likert-type rating scale, which ranged from 1 to 9. The extent of the level of sexual aversion was assessed by scores from 1 (extremely high) to 4 (extremely low), and the level of sexual arousal was from 6 (extremely low) to 9 (extremely high). No sexual response to the stimuli was given a score of 5.
Imaging
MR imaging was performed on a 1.5T scanner (GE Healthcare, Milwaukee, Wis). After acquisition of a high-resolution T1-weighted anatomic scan, subjects underwent whole-brain functional runs (echo-planar imaging [EPI], gradient-recalled echo; TR = 3000 ms, TE = 40 ms, flip angle = 90°, FOV = 240 mm, 64 × 64 matrix, twenty 5-mm axial sections) for measurement of blood oxygen level–dependent effects.
Whole-Brain Analysis
Data preprocessing and statistical analysis were performed with SPM2 (Wellcome Department of Cognitive Neurology, London, UK) running with Matlab (MathWorks, Natick, Mass). The first 4 EPI volumes were discarded to allow T1 equilibration effects. The remaining images were realigned to the first volume and normalized to the standard EPI template of the Montreal Neurological Institute (3 × 3 × 3 mm voxels). The normalized images were then spatially smoothed with a gaussian kernel at full width at half maximum of 8 mm to reduce intersubject differences in anatomy and enable the application of gaussian random field theory.
Effects at each and every voxel were estimated by using the general linear model. Voxel values for each contrast yielded a statistical parametric map of the t statistic (SPM{t}), subsequently transformed to the unit normal distribution, SPM{z}. The within-group analysis for comparisons was performed according to a random-effect model, applying 1-sample t test analysis, yielding statistical parametric maps. The resulting maps revealed strong activation and were hence thresholded at P = .001 uncorrected (spatial extent ≥10 contiguous voxels) to increase sensitivity for valence-dependent differences. Anatomical Automatic Labeling software (http://www.cyceron.fr/freeware) was applied for anatomic labeling of the extremum of an activated cluster by a set of coordinates.
Results
Behavioral Ratings
Two independent samples tests revealed a strong main effect of sexual orientation; there were significant differences between homosexual and heterosexual men in the scores of the F-F, F-M, or M-M stimuli (z = −3.369, P = .001; z = −3.836, P < .001; z = −3.847, P < .001) (Fig 1).

Mean scores of the sexual attractiveness of films showing F-F, F-M, and M-M and the maximal level of evoked sexual arousal (SA) in the 2 groups. In homosexual men, M-M stimuli induce maximal sexual arousal, whereas F-M stimuli are preferred in heterosexual men. Results of 2 independent samples test comparisons (homosexual versus heterosexual) are displayed. Blue indicates homosexual men; green, heterosexual men; the asterisk, P < .001. Error bars equal 1 SD.
The F-F and M-M stimuli induced sexual aversion in homosexual and heterosexual men, respectively, with corresponding mean scores of 3.55 ± 1.67 and 2.90 ± 1.37. The F-M and M-M stimuli were both rated as sexually arousing by the homosexual men, and the M-M stimulus (mean score = 8.30 ± 0.92) was more sexually attractive and arousing than the F-M stimulus (mean score = 5.7 ± 0.67; F = 52.000; P < .001). However, the heterosexual subjects reported the F-M stimulus (mean score = 7.90 ± 0.57) to be more sexually attractive and arousing than the F-F stimulus (mean score = 6.55 ± 1.50; F = 7.093; P < .05). There was no difference in the maximal level of sexual arousal between the homosexual and heterosexual groups (z = −1.490, P > .05).
Functional Neuroimaging
Following spatial preprocessing of the functional images, activation contrast between the conditions was estimated for each subject at each voxel by using linear regression, producing parametric t statistic maps. Sexual orientation differences in activation were assessed by using second-level 2-group unpaired t tests. The entire brain was examined for regions in which differential activity surpassed a statistical threshold of P < .001 (uncorrected) and spanned a minimum of 10 contiguous 64-mm3 voxels.
Each type of erotic film versus the rest condition induced greater activity in a large number of brain regions in both groups.
In the homosexual group, F-F versus rest, F-M versus rest, and M-M versus rest showed that some brain regions were activated in all the 3 conditions, including the left and right cerebellum, right cingulate gyrus, left middle frontal gyrus, right middle superior frontal gyrus, left middle occipital gyrus, right inferior temporal gyrus, and left temporal lobule. In within-group comparison analysis, F-F versus rest and F-M versus rest showed activation in the right cuneus, left hippocampus, and right postcentral gyrus, which was not shown in the M-M condition. Moreover, F-M versus rest and M-M versus rest showed activation in the left superior parietal gyrus; however, the right superior and inferior parietal gyri were only activated in the F-F condition. In particular, activation in the right insula and right parahippocampus was observed only in F-M versus rest, and M-M versus rest was associated particularly with activation in the left angular gyrus, left caudate nucleus, and right pallidum.
Otherwise, in the heterosexual group, F-F versus rest, F-M versus rest, and M-M versus rest showed significant activation in the left and right cerebellum, left middle frontal gyrus, left and right middle superior frontal gyri, left and right superior frontal gyri, left superior parietal gyrus, left precentral gyrus, left and right occipital lobules, left precuneus, and right middle temporal gyrus. In within-group analysis, F-F versus rest and F-M versus rest showed activation in the left anterior cingulate gyrus, right lingual gyrus, right parahippocampal gyrus, left postcentral gyrus, and left thalamus, which did not occur in the M-M condition. The right hippocampus and right precentral gyrus were consistently activated by F-M and M-M stimuli. We also found that some brain regions were activated by only 1 type of stimulus. These special neural correlates included the left angular gyrus, left hippocampus, and right thalamus for F-F versus rest; the insular cortex and vermis for F-M versus rest; and the right angular gyrus, right caudate nucleus, and right cuneus for M-M versus rest.
We compared the activation between groups during evoked sexual arousal (Tables 1 and 2). In the homosexual group, the left angular gyrus, left caudate nucleus, and right pallidum were activated by the M-M stimulus versus rest: left angular gyrus, P < .001 uncorrected, maximum at −42, −68, 48 (gyri = infinite); left caudate nucleus, P < .001 uncorrected, maximum at −8, 0, 24 (gyri = 6.25); right pallidum, P < .001 uncorrected, maximum at 14, 0, 6 (gyri = 5.83). In contrast, heterosexual men showed no activation in these regions during sexual arousal (Fig 2).
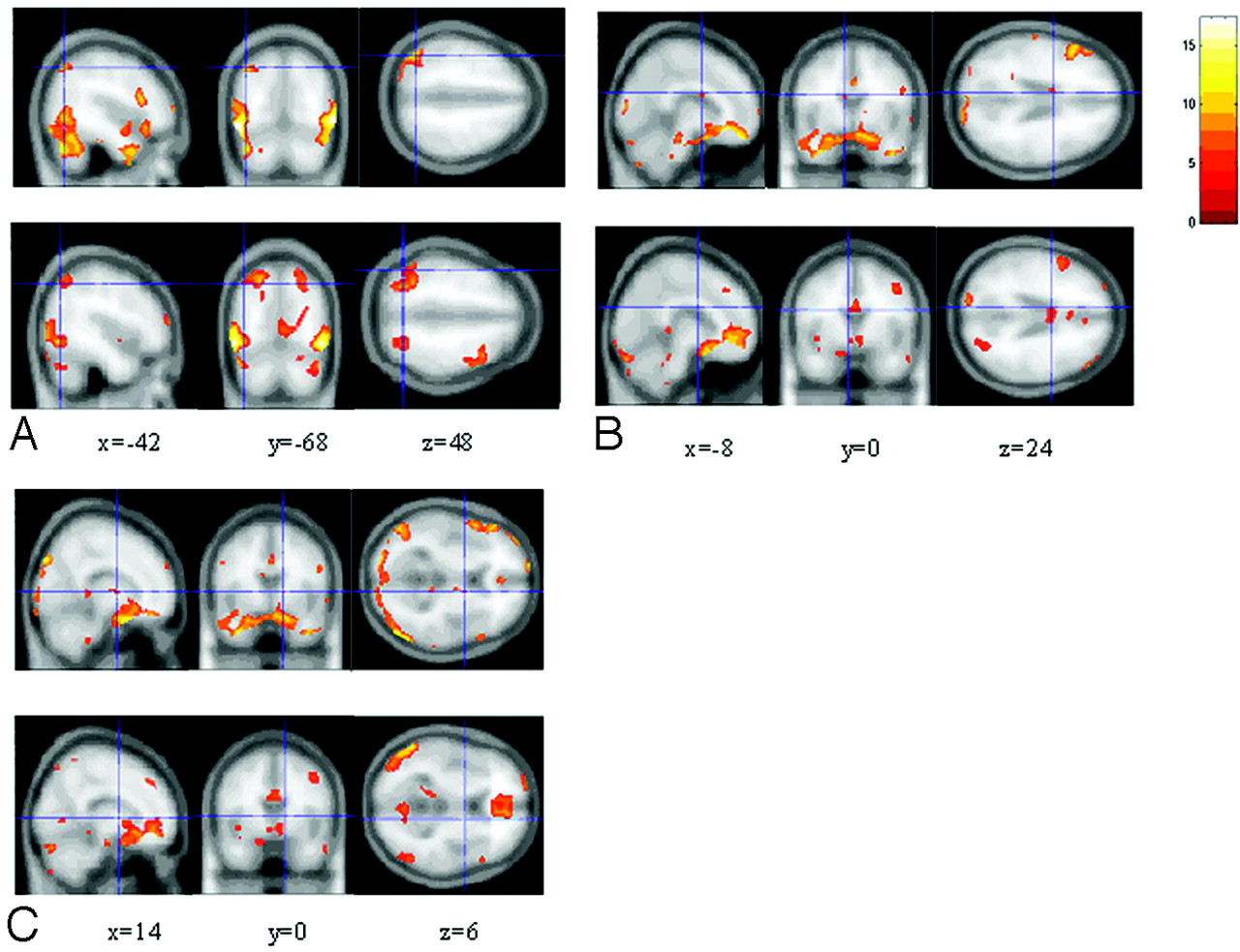
Images showing brain activation in response to M-M stimuli in the homosexual group compared with the same regions in response to F-M stimuli in the heterosexual group, by 1-sample t test analysis. In each labeled panel of 6 images, the upper rows are from homosexual men and the lower rows are from heterosexual men. Significant activation occurs in the left angular gyrus (x = −42, y = −68, z = 48) (A), left caudate (x = −8, y = 0, z = 24) (B), and right pallidum (x = 14, y = 0, z = 6) (C). The bar shows the range of the t value. Numbers in the bottom row indicate the coordinates of the Montreal Neurological Institute brain.
Brain activation of the male homosexual group during visually evoked sexual arousal in M-M versus rest condition*
Brain activation of male heterosexual group during visually evoked sexual arousal in F-M versus rest condition*
In addition, the heterosexual group showed activation in the bilateral lingual gyrus, right hippocampus, and right parahippocampal gyrus during visually evoked sexual arousal: left lingual gyrus, P < .001 uncorrected, maximum at 2, −80, −2 (gyri = infinite); right lingual gyrus, P < .001 uncorrected, maximum at 6, −70, 2 (gyri = infinite); right hippocampus, P < .001 uncorrected, maximum at 20, −36, −2 (gyri = 7.52); right parahippocampal gyrus, P < .001 uncorrected, maximum at 16, −36, −10 (gyri = 6.39). In contrast, no activation in these regions was found in homosexual men (Fig 3).

Images showing brain activation in response to F-M stimuli in the heterosexual group compared with the same regions in response to M-M stimuli in the homosexual group, by 1-sample t test analysis. In each labeled panel of 8 images, the upper rows are from homosexual men and the lower rows from heterosexual men. Significant activation occurs in the left lingual gyrus (x = 2, y = −80, z = −2) (A), right lingual gyrus (x = 6, y = −70, z = 2) (B), right hippocampus (x = 20, y = −36, z = −2) (C), and right parahippocampus (x = 16, y = −36, z = −10) (D). The bar shows the range of the t value. Numbers in the bottom row indicate the coordinates of the Montreal Neurological Institute brain.
Except for the significantly different activation in the previously mentioned regions, many regions were consistently activated in both homosexual and heterosexual men. These included the middle prefrontal cortex, bilateral temporal lobe, and postcentral gyri (P < .001); in addition, the thalamus, insula, vermis, left supramarginal gyrus, left precuneus, occipital cortex, parietal cortex, and cerebellum were activated (P < .05, ≥5 contiguous voxels).
Region-of-Interest Analysis
To compare differences in fMRI signal intensity changes across all stimulus conditions and among brain regions, we calculated the average fMRI signal-intensity change relative to the fixation baseline for each subject for regions of interest defined in the left and right amygdala and the left and right thalamus. Group-difference analysis revealed that homosexual men showed the same magnitude of activation as heterosexual men in all of these regions of interest during sexual arousal. Stimulus-condition analysis revealed that heterosexual men showed greater activation than homosexual men in the left amygdala for the M-M stimulus (z = 2.156, P < .05). However, there was no significant difference between homosexual and heterosexual men in the magnitude of activation in each region of interest for the F-F and F-M stimuli.
Analysis revealed no correlation between the magnitude of any region of interest and the mean level of sexual arousal of homosexual or heterosexual men.
Discussion
It has recently been proposed that human sexual arousal, which is usually triggered by external stimuli or endogenous factors, is a multidimensional experience comprising cognitive, emotional, motivational, and physiologic components.9 The cognitive component involves a process of appraisal through which a stimulus is evaluated as a sexual incentive. The emotional component refers to the specific hedonic quality of sexual arousal. The motivational component relates to the processes that direct behavior to a sexual goal. Finally, the physiologic component comprises the autonomic and endocrine responses (eg, cardiovascular, respiratory, and genital) associated with sexual arousal.10
Visual sexual stimuli can be viewed as emotion-laden. In humans, brain regions that are implicated in the processing of emotion-laden visual stimuli include the hypothalamus, thalamus, medial prefrontal cortex, anterior temporal cortex, occipitotemporal cortex, amygdala, hippocampal formational, and ventral striatum.11–13 Furthermore, the anterior cingulate, occipitotemporal and orbitofrontal cortices, as well as the insula, ventral striatum, claustrum, nucleus accumbens, parietal lobules, thalamus, and hypothalamus have all been shown to respond to sexually explicit films in men.9,10
Two recent studies on homosexual and heterosexual individuals provide some evidence that sexual preference has a strong impact on neuronal responses to sexually relevant stimuli. Smelling pheromonelike compounds increases neuronal activity in the medial preoptic area of the hypothalamus, and the magnitude of activation depends on sexual preference, at least in men.14 In heterosexual and homosexual men and women, male and female face stimuli elicit a stronger activation of the dorsomedial thalamus and medial orbitofrontal cortex if the sex of the face matches the sexual preference of the observer.15
The most important result of the present study was that in homosexual men, the neuroanatomic correlates of visually evoked sexual arousal were different from those in heterosexual men. This is consistent with our hypothesis that different neural circuits exist in homosexual and heterosexual men.
Hypersexuality has been reported in patients with lesions localized to the head of the caudate.16 Moreover, activation of the right caudate nucleus is correlated with the intensity of the urge to perform handwashing rituals or the intensity of various provoked obsessive-compulsive disorder (OCD) symptoms.17,18 In both the sexually arousing condition and in the OCD symptom-provoking paradigm, subjects are simultaneously confronted with the urge to act and with the practical impossibility of doing so, and the 3 regions—orbitofrontal cortex, cingulate cortex, and caudate nuclei—are known to be part of a circuit, also including the thalamus, that may be involved in the regulation of both consciously perceived emotional concerns and repetitive behavior.18 However, the results of the present study, that the caudate nuclei of heterosexual men were not activated, is not consistent with the findings of Rauch et al.18 In contrast, in their study, this anatomic structure was activated in homosexual men during sexual arousal. The reason for this difference is not known. Concerning the responses of subjects stimulated by erotic film excerpts, we consider that they did not have a conscious struggle with obsession similar to patients with OCD, and the cause of activation of the caudate nuclei in response to erotic stimuli was not same as that of patients with OCD. We presume that the caudate nuclei of homosexual men have a different function in the regulation of perceived emotional concerns than those of heterosexual men.
The pallidum belongs to the paralimbic system and functions in evaluating internal motivation as well as extrinsic stimuli and in regulating dependent behaviors. When heterosexual women view an erotic film, several paralimbic areas, including the globus pallidus, inferior frontal cortex, insular gyrus, cingulate gyrus, and thalamus are activated, along with sexual stimulation.19 However, the study of Stoléru et al9 showed no activation in the pallidum of healthy heterosexual men during sexual arousal. These studies demonstrated that there is a sex difference in activation of the pallidum during human sexual arousal. Using the processing-mode hypothesis, Hamann et al8 analyzed the sex differences in brain activation in response to sexual stimuli. This hypothesis predicts that men show greater brain activation than women in specific regions after controlling for arousal. Possibly, the reason for different activation in the pallidum between homosexual and heterosexual men is that there are different neural pathways in the 2 groups, according to the processing-mode hypothesis.
Another important result of the present study was that the lingual gyrus, hippocampus, and parahippocampal gyrus were activated in heterosexual men but not in homosexual men.
Some studies have identified sex differences in mental rotation tests. That activation of the lingual gyrus in heterosexual men is greater than that of heterosexual women during egocentric mental rotation and demonstrates that men have more extensive semantic or earlier visual processing.20 Several independent studies consistently demonstrated low scores (female-typical) by homosexual men in basic spatial ability tests (such as mental rotation and spatial perception) compared with heterosexual men.21 The possible reason for the absence of activation in the lingual gyrus of homosexual men is that they have a lower ability in spatial perception than heterosexual men.
The hippocampus is an important structure that has long been known to play a key role in memory and learning, and some studies have shown that the hippocampus exhibits sexual dimorphism at the cellular level. There is evidence that testosterone plays a role in the dimorphism of the hippocampus.22 Animal studies have demonstrated that neonatal males have more granule cells than females in both the dorsal and ventral dentate gyri and a greater volume in all layers.23 An fMRI study revealed sex-related differences in the lateralization of hippocampal activation and cognitive strategy.24 In the present study, there was significant activation in the hippocampus of heterosexual men responding to visual sexual stimuli. In contrast, we did not find activation in the hippocampus of homosexual men during sexual arousal. This interesting result suggests that there are sexual orientation–related differences in emotional memory and cognitive strategy.
The amygdala is an important structure in modulating emotion. Animal experiments have demonstrated that focal damage of the amygdala induces a constellation of symptoms including loss of fear and anger, increased exploratory behavior, hyperesthesia, and bulimia.25 An fMRI study reported that the amygdala is not activated in healthy men in response to sexually arousing stimuli, and a PET study reported that only a cluster of deactivation occurs in parts of the amygdala during human male ejaculation.26 In other neuroimaging studies, decreased amygdala activity was found when volunteers who were deeply in love viewed pictures of their loved ones (Bartles et al, 200027). Breiter et al28 also showed a similar deactivation when volunteers experienced a cocaine rush, and they suggested that amygdala deactivation is correlated with euphoric psychological states. In the present study, we found no activation in the amygdala of homosexual or heterosexual men. However, some fMRI studies have reported that the amygdala is more strongly activated in men than in women in response to sexually arousing stimuli.7,8 Given these inconsistent results, we cannot draw a final conclusion about the function of the amygdala during human male sexual arousal.
We found some areas of activation in the same brain regions of both groups; these included the thalamus, insula, and cerebellum.
In primates, it has been suggested that the thalamus is part of a neural network including the amygdala and orbitofrontal cortex.29 The thalamus represents a hub through which any area in the cortex can communicate with any other area. This extensive thalamocortical interconnectivity has been theorized to constitute a neuronal basis for conscious awareness and is implicated in the cognitive dimension of sexual arousal.30 We suggest that the cognitive function of the male thalamus is not affected by sexual orientation.
Intraoperative stimulation of the insular cortex before temporal lobectomy in patients with epilepsy evokes autonomic reactions.31 This is consistent with the fact that the insular cortex is highly interconnected with regions involved in autonomic regulation.32 Given such findings, the insular activation in both groups might be a neural correlate of the autonomic changes associated with sexual arousal.
Although the cerebellum is essentially involved in motor control, some studies indicate that it also contributes to cognitive33 as well as to emotional and motivational processes.34–36 Our results are consistent with reports of cerebellar activation during the heroin rush,37 listening to pleasant music,38 and monetary rewards.39 Similarly, cerebellar activation has been found in response to excerpts from erotic films.40,41 Moreover, clinical studies have revealed that large lesions of the cerebellar vermis in humans may cause blunting of affect and inappropriate behavior.42,43 Thus, possible interpretations of the cerebellar activation we observed are that they might be related to the emotional, motivational, and/or motor imagery processes induced by sexual stimuli.
Conclusion
In this study, we investigated brain activation of homosexual and heterosexual men in response to sexual stimuli, and we demonstrated male sexual orientation differences. By providing evidence for differences in brain activation, these results point to the existence of differences in the way homosexual and heterosexual men process visual sexual stimuli.
Acknowledgments
We thank Drs Lain Bruce, Roger A. Gorski, and Xiao-jun Xu for their judicious comments. We are also grateful to Dr Peng Li and Fei-yan Chen for their help regarding statistical analyses. Finally, we wish to thank the staff of the Centre of Neuroimaging, the First Affiliated Hospital, College of Medicine, Zhejiang University, for their skillful technical assistance.
References
- Received May 16, 2008.
- Accepted after revision June 26, 2008.
- Copyright © American Society of Neuroradiology





{kind=link}
{kind=link}
{kind=link}