
Abstract
BACKGROUND AND PURPOSE: MR spectroscopy allows the noninvasive evaluation of in vivo brain metabolites. Our purpose was to use this technique to assess metabolic alterations in the human cerebrum during growth, maturation, and aging.
METHODS: Ninety normal human brains in subjects aged 4 to 88 years were examined with multivoxel proton MR spectroscopy. Spectra were obtained from specific voxels of 2.5 cm3 in the gray and white matter of the centrum semiovale. The ratios of N-acetylaspartate (NAA) to choline (Cho) were calculated to describe age-dependent alterations in cerebral metabolites.
RESULTS: White matter NAA/Cho ratios showed rapid growth during the first decade and reached a maximum value in the second or early third decade, followed by a steady decline starting in the latter half of the third decade. The maximum peak ages for NAA/Cho were 21.9, 17.6, and 15.9 years (mean, 18.5 years) for the anterior, middle, and posterior white matter, respectively. A significant cerebral laterality of the white matter NAA/Cho was found in male subjects during development. The growth spurt and age-related decline of the white matter NAA/Cho were steeper in male than in female subjects. In contrast, the gray matter NAA/Cho showed a gradual decline with age.
CONCLUSION: Proton MR spectroscopy shows significant regional and sex differences in the level of cerebral metabolites during the process of growth, maturation, and aging. This technique may play an important role in clinical applications for various conditions of metabolic disorders of the human brain.
Although the macroscopic and microscopic evolution and involution of the human cerebrum have been studied in detail (1–5), little is known about which neuronal components increase during the growth period and decrease later in life. With the advent of MR imaging, it has been possible to follow the steps of development and aging of the human brain in vivo (6–9). However, the detailed process of age-dependent alterations of the brain has not yet been fully analyzed with MR imaging.
Recent innovations in MR technology have been established in the field of spectroscopy (10–12). Many investigators have used proton MR spectroscopy to study brain metabolism in vivo; in particular, the changes that occur in specific chemical components of the human brain in normal and pathologic states (12–24). Several studies have focused on the development or aging of the human brain, using primarily single-voxel MR spectroscopy (25–31). To our knowledge, no report has fully described the regional and sex differences in metabolites of the human brain in vivo across a wide range of ages. We thought that because of its superior sensitivity to minute metabolic composition, MR spectroscopy would be especially useful for the evaluation of alterations in neuronal metabolites that occur between childhood and old age.
The purpose of this study was to develop a proton MR spectroscopic database of the normal human brain in vivo, with particular emphasis on the regional and sex differences in cerebral metabolites during the process of growth, maturation, and aging.
Methods
Subjects
Between October 1996 and June 1998, a total of 90 healthy volunteers underwent proton MR spectroscopy of the brain. The subjects included five male and five female subjects for each age decade (total, 45 males and 45 females) with ages ranging from 4 to 88 years (mean, 45.6 years). All examinations were performed with the informed consent of the subjects or their parents, and were in compliance with institutional guidelines. None of the subjects had a history of psychiatric or neurologic disorders and none had intracranial pathologic lesions revealed by T1- and T2-weighted studies obtained at the time of MR spectroscopy. No sedation was used for any subject.
MR Study
Proton spin-echo MR imaging and spectroscopy were performed on a 1.5-T scanner using the standard quadrature head coil and actively shielded gradients. Spin-echo T1-weighted (540/14/2 [TR/TE/excitations]) and fast spin-echo T2-weighted (5400/99eff/2; echo train length = 11) sequences were used to obtain images covering the whole brain.
Multivoxel proton MR spectroscopic data sets were acquired by using point-resolved spectroscopy with acquisition parameters of 1500/270/2 and a transverse field of view of 18 × 18 cm2 with a 16-in × 16-in matrix. After three orthogonal base images were obtained with automatic shimming of the magnetic field, a 20-mm-thick volume of interest (VOI) was identified, resulting in an effective voxel size of 2.5 cm3, which was consistently located in the centrum semiovale (ie, parallel to the planum sphenoidale and immediately above the body of the corpus callosum). The local magnetic homogeneity within the VOI was further optimized by an additional manual shimming so as to give a maximum proton signal from water. Usually, a half-height line width of the water signal of 0.15 ppm was achieved. Starting with shim settings that were known to result in a near-optimal water signal, additional manual shimming took 2 to 5 minutes. MR spectroscopic data were accumulated after the optimal water signal was suppressed by the chemical shift-selective technique. The whole examination, including MR imaging and spectroscopy, could be completed within 30 minutes.
Using fast Fourier transformation of the acquired data sets, we obtained spectra from 12 specific voxels within the VOI outlined on the MR images. Voxels were selected in the following manner: six areas were located in bilateral parasagittal anterior, middle, and posterior cortices, characterized primarily by frontal, paracentral, and parietal mesial gray matter, and six were in lateral anterior, middle, and posterior regions containing predominantly frontal, precentral, and parietal white matter, respectively. The six lateral voxels were selected so as to contain as much white matter as possible, avoiding mesial and lateral cortical sulci and gray matter. Typical locations of the analyzed voxels are shown in Figure 1A and B (page 129). After manual baseline and linear phase corrections, the spectra were quantified by means of gaussian curve fittings to measure the integrated areas under the peaks. Three dominant spectra were analyzed: choline-containing compounds (Cho) at 3.21 ppm, creatine/phosphocreatine (Cr) at 3.04 ppm, and N-acetylaspartate (NAA) at 2.02 ppm.

A and B, Sagittal (A) and axial (B) MR images of a healthy 20-year-old woman show the outlines of the spectroscopic VOI and typical position of 12 voxels in the centrum semiovale. Six voxels were in bilateral anterior, middle, and posterior regions containing predominantly white matter (white open boxes) and six voxels were in bilateral mesial cortex with mostly mesial gray matter of the anterior, middle, and posterior regions (black open boxes), respectively.
C, Proton spectra obtained in right middle white matter (white open box) are shown. NAA, Cho, and Cr peaks were quantified by applying gaussian curve fittings.
Data Analysis
Our analysis of the data included calculating the ratios of the integrated areas under the peak and then plotting the ratios with age. From these quantified peak areas, we calculated the ratios of NAA/Cho. To describe alterations in the ratios with age, a least-squares analysis was performed using KaleidaGraph software (Synergy, Reading, PA) to adjust parameters of model functions chosen on a heuristic basis for reasons of simplicity and plausibility. These consisted of both an exponential development part and a linear regression part:  where R(t) is the ratio of spectral peak areas at the age of t years, and a, b, c, and d are parameters influencing the level of the curve and the extent of increase or decrease. Then, the maximum peak age (tmax) was obtained at the age when the derivative of the function R(t) with respect to t was zero if present, as follows:
where R(t) is the ratio of spectral peak areas at the age of t years, and a, b, c, and d are parameters influencing the level of the curve and the extent of increase or decrease. Then, the maximum peak age (tmax) was obtained at the age when the derivative of the function R(t) with respect to t was zero if present, as follows:  For statistical analysis, Student's t test and analysis of variance were performed using StatView (Abacus Concepts, Berkeley, CA).
For statistical analysis, Student's t test and analysis of variance were performed using StatView (Abacus Concepts, Berkeley, CA).
Results
Representative proton spectra and peak fittings in a healthy 20-year-old woman are shown in Fig 1C. The spectral peaks of NAA, Cho, and Cr were sharp and reasonably separated. Scatter plots and least-squares analyses for the NAA/Cho ratios are shown in Figures 2 and 3.

A and B, Scatter plots and fitting curves of averaged NAA/Cho ratios as a function of age in white matter (A) and gray matter (B) of the centrum semiovale.
Differences in the NAA/Cho ratios with age were found between gray and white matter; among anterior, middle, and posterior regions; and between right and left cerebra. Significant differences in the ratios were also observed between male and female subjects.
Regional Differences in Age-dependent NAA/Cho Ratios
Differences in NAA/Cho ratios for anterior, middle, and posterior white matter were significant (P < .001). The age-dependent NAA/Cho ratios in white matter showed rapid growth during the first decade and reached their maximum value in the second or early third decade (Fig 2A). The growth spurt of NAA/Cho in the posterior white matter was steeper than that in the anterior and middle white matter. The averaged NAA/Cho ratios for bilateral white matter reached a maximum at the ages of 21.9, 17.6, and 15.9 years in the anterior, middle, and posterior white matter, respectively. The mean age for the maximum NAA/Cho value in the three regions of white matter was 18.5 years.
Cerebral lateralization was observed in the NAA/Cho ratio in homologous areas of white matter during development. A higher level of NAA/Cho was found in right-sided white matter relative to that of left-sided white matter in the first and second decades (Fig 3A–C). A significant laterality of the NAA/Cho ratio was found in the posterior white matter during the first decade of life (P = .019). The maximum NAA/Cho ratio occurred 1.1 to 4.0 years earlier in right-sided white matter than in left-sided white matter.
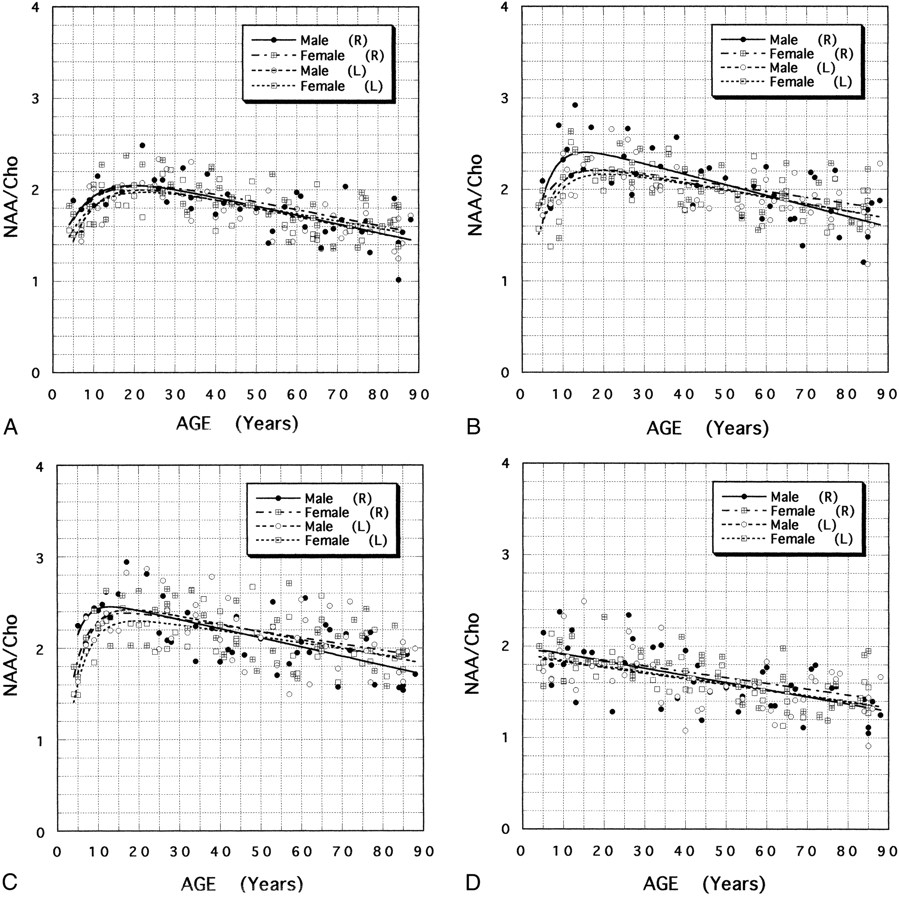
A–F, Scatter plots and fitting curves for male and female NAA/Cho ratios as a function of age in anterior (A), middle (B), and posterior (C) white matter, and anterior (D), middle (E), and posterior (F) gray matter of the right (R) and left (L) centrum semiovale.

Continued.
There was a steady, almost linear, decrease in the white matter NAA/Cho ratio, starting in the late third decade and continuing into old age. In comparing 85-year-old with 18.5-year-old subjects, we found a 24.6%, 23.0%, and 21.6% reduction in the averaged NAA/Cho for bilateral anterior, middle, and posterior white matter, respectively.
Differences in the NAA/Cho ratios for anterior, middle, and posterior gray matter were significant (P < .001). Gray matter NAA/Cho ratios showed a gradual decrease with age from childhood to old age (Fig 2B). The age dependency of NAA/Cho was almost negatively linear, and the reduction rates in the averaged NAA/Cho ratios between 18.5-year-old and 85-year-old subjects were 24.5%, 21.2%, and 22.0% for bilateral anterior, middle, and posterior gray matter, respectively.
Sex Differences in the Age-dependent NAA/Cho Ratio
Significant differences in NAA/Cho were found between male and female subjects during development and aging. The growth spurt of the white matter NAA/Cho was steeper in male than in female subjects. The maximum NAA/Cho value in age-dependent alterations of white matter occurred 1.4 to 3.2 years earlier in male than in female subjects.
Notable sex differences were found in the degree of cerebral laterality in NAA/Cho ratios. NAA/Cho levels were higher in right-sided white matter in males in their first and second decades (Fig 3A–C), with a significant laterality in the NAA/Cho ratio in the posterior white matter in male subjects during the second decade (P = .011). No significant laterality was observed in NAA/Cho ratios in female subjects during development, which were almost the same as those in the left-sided white matter of male subjects.
From the latter half of the third decade, the involutional process of the white matter NAA/Cho ratio was almost linear, but there was a sex difference in the reduction rate of NAA/Cho. The decline rate of the right-sided white matter NAA/Cho ratio in male subjects was greater than on the left side and greater than the decline rate bilaterally in female subjects.
The decline rate of the gray matter NAA/Cho was almost linear with age; and, in male subjects, it was greater on the right side than on the left side and greater than the decline rate bilaterally in female subjects (Fig 3D–F).
Discussion
Proton MR spectroscopy is a useful in vivo examination for analyzing the metabolites of the human brain in small concentrations (12–14). However, there are shortcomings with the practical application of proton MR spectroscopy in the reliable assessment of the metabolic composition of the brain, because of inaccuracies in quantification caused by magnetic inhomogeneities and because of degradation of the spectral resolution from the overlapping of peaks from unwanted metabolites. We made several adjustments in an attempt to overcome these shortcomings.
First, the most homogeneous magnetic field available was secured over a large VOI located in the centrum semiovale by automatic and manual shimming techniques to attain the highest detectability of proton signals from water that were suppressed thereafter. Second, we used a long TE of 270 milliseconds. A long TE has the advantage of providing a spectrum dominated by the major peaks of interest, with negligible contributions from chemical composites with short T2, such as fat and macromolecules. Third, the use of multivoxel proton MR spectroscopy provided better signal-to-noise ratio, since it is possible to define signal sources from a wide area of the brain simultaneously. As a consequence, three dominant spectra of NAA, Cho, and Cr were sharp and reasonably determined with superior spectral resolution (Fig 1C).
NAA is an amino acid located exclusively in neuronal cell bodies, dendrites, and axons (32, 33). Previous studies have reported that the NAA level can be altered by neuronal cell death or other neuronal damage in gray matter (16, 23), whereas in white matter, lower NAA has been interpreted to reflect diffuse axonal loss or damage (14–20). In contrast, the Cho level has been regarded as a marker of cellular density, since Cho is the precursor for phosphatidylcholine, which is a major constituent of the cell membrane (34, 35). Thus, the NAA/Cho ratio is regarded as a significant indicator in the assessment of neuronal activities, because it represents a relative ratio of neuronal density to cellular density. In practice, alterations in the NAA/Cho ratio have been reported to result in findings suggestive of neuronal damage that are consistent with pathologic findings of neuronal disorders (15, 16, 18–21, 23). The NAA/Cr ratio has occasionally been used to examine metabolic abnormalities (14–22); however, the importance of the peak consisting of Cr is not as explicit as that of Cho. According to Möller et al (21), the most sensitive marker for abnormalities in the cerebral metabolism is NAA/Cho, which was found to decrease significantly in AIDS patients with CNS involvement.
One of the most significant findings of the present study was the age dependency of NAA/Cho ratios in different gray and white matter structures. The gray matter NAA/Cho ratios decreased gradually from childhood to old age, whereas the white matter NAA/Cho ratios showed a growth spurt during the first decade and reached maximum values in the second or early third decade. We suspect that the elevation in white matter NAA/Cho must be related to the increase in neuronal content relative to the density of axons, which may correspond to neuronal maturation.
Because the increase in the number of neuronal cells reaches its peak by about 6 months after birth, maturational changes of the centrum semiovale after infancy occur through significant increases in the number and size of axonal and dendritic fluctuations (1). Major events in the maturation of the cerebrum take place during the first few years of life (2), but the myelination that is most closely related to full functional capacity of neuronal interconnections is not completed before adolescence, a period during which the maturational process is associated with significant alterations in the biochemical composition of the cerebrum (1).
In our study, the maximum peak NAA/Cho ratios in white matter occurred at the age of 21.9 years for the frontal association area, 17.6 years for the precentral motor area, and 15.9 years for the parietal sensory association area. The averaged NAA/Cho ratios in these various white matter areas reached a peak at the age of 18.5 years. These ages may correspond to fully developed axonal processes with myelination; that is, the maturation of the white matter of the centrum semiovale. Our results suggest that the white matter of the cerebrum matures in a dorsal to rostral direction, which corresponds to the order of myelination in the subcortical white matter elucidated in anatomic studies (2). In addition, cerebral lateralization was observed in NAA/Cho of the white matter in the process of maturation. Cerebral white matter may mature 1.1 to 4.0 years faster on the right side than on the left side.
Beginning late in the third decade, a gradual but steady decrease in NAA/Cho takes place in the white matter. The age-dependent decrease in white matter NAA/Cho may correspond to a deterioration of some of the physiological functions that show a slow, nearly linear, decline starting at 30 years of age and progressing into late maturity (4), such as a steady decline in cognitive function. We found a 17% to 30% reduction in the amount of white matter NAA/Cho in 85-year-old subjects relative to that in younger adults, which is consistent with the finding that, in elderly adults, the activity of certain enzymes or the level of selected proteins or RNA molecules ranges 5% to 30% below that in young adults (5).
Another important finding of our study was the significant difference between male and female subjects in white matter NAA/Cho ratios during development and aging. The maximum value of white matter NAA/Cho occurred 1.4 to 3.2 years earlier in male than in female subjects. Moreover, the rate of decline of this ratio was steeper in male than in female subjects. The most interesting finding was the significant cerebral laterality in NAA/Cho ratios that occurs primarily in male subjects during a particular period of development, in contrast to the almost identical bilateral age-dependent alterations in NAA/Cho that occur in female subjects. Our results suggest that the sex differences in the cerebra are related to the asymmetry of the age-dependent alterations in NAA/Cho in male subjects, especially in the parietal lobe. It has been postulated, on the basis of the sexual dimorphism in the corpus callosum, that the male brain is more asymmetric than the female brain (36, 37). The evidence suggests that men's and women's brains are organized along different lines from early in life, and that sex hormones play roles in such differentiation (38, 39). Our present findings suggest that the same hormonal factors influence the functional potential of the right cerebral hemisphere in male children, especially in the parietal lobe.
Our study has several limitations. First, although we investigated a wide range of subjects stratified by age between 4 and 88 years, we did not include subjects in the important first four years of life, which is a critical period for the developing brain. For this reason we could not assess the growth process completely. We chose not to include children in this age group because it is not possible to obtain reliable spectra without sedating infants, and the effects of medication on the spectra are not clear.
A second limitation is that the signal for each of the voxels did not come entirely from gray or white matter, owing to a voxel size on the order of 2.5 cm3. We tried to select voxels that contained as much gray or white matter as possible while avoiding other tissue; however, it is possible that the observed level in the gray matter NAA/Cho ratios was influenced by white matter signal. It would have been desirable to use a voxel size smaller than 1 cm3, but that is not yet easily attainable on our imager.
The third limitation is the lack of absolute quantitation of cerebral metabolites. However, there may be inaccuracies in absolute quantification due to magnetic inhomogeneities. In addition, the absolute peak areas of each chemical metabolite must be affected by inherent partial volume effects of various tissues within the VOI and possible radiofrequency instability among analyzed voxels. On the other hand, the ratio of peak areas is considered to be an appropriate marker for evaluating the relative alterations in the metabolites, because of a certain cancellation of the larger portion of errors caused by contaminated tissues and instrumental instability. For instance, although CSF within the voxels chosen for analysis may not contain significant amounts of the metabolite measured by proton MR spectroscopy, the variable contribution of CSF to the voxel volume should not affect metabolic ratios. For a primary data analysis, it may be sufficient to determine peak ratios stratified by age between 4 and 88 years of life.
Conclusion
Our findings indicate that proton MR spectroscopy can detect regional and sex differences during the process of growth, maturation, and aging of the human cerebrum in vivo. We observed significant age-dependent alterations in NAA/Cho ratios between the ages of 4 to 88 years in the gray and white matter, the right and left cerebra, and the anterior, middle, and posterior regions of the centrum semiovale. Significant sex differences were also found in the degree of cerebral laterality of the NAA/Cho ratios with age. The data may serve as a basis for studies of normal human neurochemistry as well as of metabolic alterations in diseases of the cerebrum.
Acknowledgments
We thank Katsuya Maruyama for technical assistance. We are also grateful to Yutaka Kuru, Toyomi Shirakawa, and Norio Iriguchi for their helpful discussions and comments on the manuscript.
Footnotes
1 Address reprint requests to Tsuyoshi Kadota, MD, PhD.
References
- Received November 19, 1999.
- Accepted after revision June 14, 2000.
- Copyright © American Society of Neuroradiology
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Changes in brain metabolite levels across childhood
- Evaluation of Normal Changes in Pons Metabolites due to Aging Using Turbo Spectroscopic Imaging
- Conversion of Ischemic Brain Tissue Into Infarction Increases With Age
- NMR Spectroscopy and Pediatric Brain Tumors
- Altered neurometabolite development in HIV-infected children: Correlation with neuropsychological tests