
Abstract
BACKGROUND AND PURPOSE: Experienced freedivers can endure prolonged breath-holds despite severe hypoxemia and are therefore ideal subjects to study apnea-induced cerebrovascular reactivity. This multiparametric study investigated CBF, the spatial coefficient of variation as a correlate of arterial transit time and brain metabolism, dynamics during prolonged apnea.
MATERIALS AND METHODS: Fifteen male freedivers (age range, 20–64 years; cumulative previous prolonged breath-holds >2 minutes and 30 seconds: 4–79,200) underwent repetitive 3T pseudocontinuous arterial spin-labeling and 31P-/1H-MR spectroscopy before, during, and after a 5-minute breath-hold (split into early and late phases) and gave temporally matching venous blood gas samples. Correlation of temporal and regional cerebrovascular reactivity to blood gases and cumulative previous breath-holds of >2 minutes and 30 seconds in a lifetime was assessed.
RESULTS: The spatial coefficient of variation of CBF (by arterial spin-labeling) decreased during the early breath-hold phase (−30.0%, P = .002), whereas CBF remained almost stable during this phase and increased in the late phase (+51.8%, P = .001). CBF differed between the anterior and the posterior circulation during all phases (eg, during late breath-hold: MCA, 57.3 ± 14.2 versus posterior cerebral artery, 42.7 ± 10.8 mL/100 g/min; P = .001). There was an association between breath-hold experience and lower CBF (1000 previous breath-holds reduced WM CBF by 0.6 mL/100 g/min; 95% CI, 0.15–1.1 mL/100 g/min; P = .01). While breath-hold caused peripheral lactate rise (+18.5%) and hypoxemia (oxygen saturation, −24.0%), cerebral lactate and adenosine diphosphate remained within physiologic ranges despite early signs of oxidative stress [−6.4% phosphocreatine / (adenosine triphosphate + adenosine diphosphate); P = .02].
CONCLUSIONS: This study revealed that the cerebral energy metabolism of trained freedivers withstands severe hypoxic hypercarbia in prolonged breath-hold due to a complex cerebrovascular hemodynamic response.
ABBREVIATIONS:
- ATP
- adenosine triphosphate
- ASL
- arterial spin-labeling
- ASL-sCoV
- spatial coefficient of variation of CBF (by ASL)
- ATT
- arterial transit time
- CVR
- cerebrovascular reactivity
- HR
- heart rate
- Pi
- inorganic phosphates
- PCr
- phosphocreatine
- pO2
- partial pressure of oxygen
- SpO2
- oxygen saturation
Freedivers acquire the ability to voluntarily breath-hold for several minutes: One breath-hold can be extended up to the world record of 11 minutes and 30 seconds.1 However, animal studies have suggested that threshold times for hypoxia-induced neuronal cell death can be as low as 2 minutes and 30 seconds.2 The regulation and sufficiency of cerebrovascular reactivity (CVR) to prevent ischemic brain damage during prolonged breath-hold are unclear. Such knowledge may hold clues to explain patterns of brain damage in adverse diving outcomes and may provide suggestions for targeted therapies. It can also be useful to deduce CVR in medical conditions of repetitive hypoxemia such as obstructive sleep apnea syndrome.3
Previous MR imaging breath-hold experiments involved hypercarbic air composition or the use of relatively short breath-holds.4 There are, however, crucial differences between experimental setups applying altered gas composition and those using voluntary breath-hold. In the latter, the human body can be examined as a closed system of internal metabolic gas exchange, while altered-ventilation gas studies are an “open system” approach with ventilation at variable frequencies as a complicating factor.5 Quite different from short breath-holds, prolonged breath-hold for several minutes is furthermore a unique mental challenge demanding an emotional preparation, for which freedivers need to train and which may have an independent effect on CVR.6 The putative effect of experience from previous prolonged breath-holds on CBF must be mentioned in this context and has not been explored.
Arterial spin-labeling (ASL) perfusion MR imaging allows an in vivo assessment of absolute CBF both regionally and on a vessel-selective level. ASL studies already successfully identified chronic CVR alterations due to obstructive sleep apnea as well as acute alterations during very short breath-holds.5,7 A corollary ASL measure is the spatial coefficient of variation (ASL-sCoV) of the CBF image itself, which was recently identified as a correlate for arterial transit time (ATT) and is a useful alternative when direct ATT estimations are not feasible.8 Thus, ASL seems a promising technique to study CVR in prolonged breath-hold.
Blood gas alterations trigger a cerebrovascular but also a peripheral vascular reaction in the body. During breath-hold, for example, the CBF increases and hence faster O2 supply facilitates the maintenance of an aerobic brain metabolism under hypoxemia—that is, to prevent the depletion of adenosine triphosphate (ATP) and the accumulation of lactate from anaerobic glycolysis. The sufficiency of CVR can be studied indirectly by 31P-MR spectroscopy, which provides estimates of ATP and related metabolites,9 as well as by 1H-MR spectroscopy, which may show lactate if the metabolite is elevated beyond physiologic ranges.
The goal of the present study in experienced freedivers was to explore the following: 1) acute CVR during a prolonged breath-hold challenge with ASL, and 2) the sufficiency of CVR to maintain aerobic cerebral energy metabolism as measured by 31P- and 1H-MR spectroscopy. In addition, this study investigated whether pre-existing experience with prolonged breath-holds has an influence on the CVR during the breath-hold experiment.
Materials and Methods
Participants
Fifteen experienced male freedivers (median age, 36.0 years; 95% CI, 32.0–50.0 years) participated (Table). Inclusion criteria were adult age and an ability to breath-hold for >4 minutes without stress symptoms (ie, tachycardia, oxygen saturation [SpO2] below 60%, delirium). Exclusion criteria were pre-existing cardiac or neurologic disorders and current smoking. The participant's age and cumulative number of breath-holds longer than 2 minutes and 30 seconds during the volunteer's lifetime (estimated by interview report) were registered as potential influential factors.
Anthropometric data of freediver volunteersa
Study Design
Preparatory evaluations involved questionnaires regarding claustrophobia, noise tolerance. or issues with restraints. All participants were instructed to refrain from meals and caffeine for at least 2 hours before MR imaging to reduce perfusion confounders.10 Preparation (eg, meditation, test placement on the MR imaging table) was allowed. MR imaging–electrocardiography and finger oximetry guaranteed continuous monitoring of SpO2 and heart rate (HR). Arterial CO2 measurements were not permitted by the hospital ethics committee, but venous blood gas was repetitively analyzed. Continuity of breath-hold and consciousness were visually monitored (L.E.) to guarantee safety and correct measurements.
The MR imaging protocol consisted of a 3D T1-weighted acquisition followed by 5 ASL scans. Participants were instructed to breathe with normal frequency to obtain the baseline CBF (baseline phase). Hyperventilation to reduce blood CO2 or increase O2 concentrations was a forbidden confounder. The participants gave an acoustic signal when starting the 5-minute breath-hold, when 2 consecutive ASL scans were acquired (early and late breath-hold phases). After the second breath-hold scan, the participant was instructed to breathe again at normal frequency (approximately 16/min.). Without delay, 2 normal-breathing ASL scans (early and late recovery phases) were obtained. A subset of participants was available for 31P- and 1H-MR spectroscopy (n = 11 and 8 participants, respectively) during separate sessions to determine relative brain metabolites and, again, venous blood gas. The study was approved by the University Hospital Bonn, Germany ethics committee. All participants provided written informed consent.
MR Imaging Sequences
All imaging was performed on a 3T Ingenia MR imaging scanner (Philips Healthcare, Best, the Netherlands). The T1-weighted MPRAGE sequence was 1 × 1 × 1 mm3. The 5 identical pseudocontinuous ASL sequences were acquired with a 3D gradient- and spin-echo readout (5 segments; acquisition voxel size, 3.75 × 3.75 × 6 mm3; FOV, 240 × 240 × 96 mm3; TE/TR, 8/4.28 s; labeling duration, 1.8 s; radiofrequency labeling pulse duration/interval, 0.7/1.41 ms; postlabeling delay, 2 s; 4 background suppression pulses; scan time, 2 minutes and 30 seconds per scan phase). Each sequence consisted of 2 M0 images and 2 control-label pairs. A labeling distance of 13 cm (middle slice of the ASL stack to labeling plane) was chosen.
Image Processing
Image processing was performed with ExploreASL (www.ExploreASL.com), a toolbox based on SPM12 routines (http://www.fil.ion.ucl.ac.uk/spm/software/spm12), which was initiated through the European Union–funded European Cooperation in Science and Technology (COST) Action ASL In Dementia (http://s434060124.online.de/aslindementiacms/), aiming at harmonizing ASL image processing for single- and multicenter ASL studies.11 Image-processing steps were the following: automated segmentation of 3D T1-weighted images using the Computational Anatomy Toolbox (CAT12 toolbox) rigid-body registration of CBF to the gray matter partial volume map and spatial normalization into a common space using the Diffeomorphic Anatomical Registration Through Exponentiated Lie Algebra Toolbox (DARTEL, part of SPM).12 M0 images were masked, iteratively smoothed, and extrapolated outside the mask, and CBF was quantified using a single-compartment quantification model.13 The ROIs analyzed were the total (cortical) GM and the total white matter. The GM was subsegmented into vascular territories of the anterior, middle, and posterior cerebral arteries. The vascular territories were delineated in the common space on the Montreal Neurological Institute atlas according to Tatu et al.14
The ASL-sCoV was defined as the SD of CBF divided by the mean CBF, within the total cortical GM (gray matter probability > 0.7)8:
 Partial volume effects within the GM and WM ROIs were corrected using the method of Asllani et al.15
Partial volume effects within the GM and WM ROIs were corrected using the method of Asllani et al.15
MR Spectroscopy
Proton-decoupled 31P spectra were acquired with a dual-tunable 31P/1H birdcage transmit/receive head coil (1024 data points; 3-kHz sampling; TR, 4 seconds; 4 signal averages) as a continuous time-series with 16-second duration for each spectrum, comprising a total of 40 scans before, during, and directly after breath-hold in a 25-mm-thick axial slice on the level of the basal ganglia. 31P signals were processed by the AMARES algorithm of the JAVA-MRUI software (http://www.jmrui.eu/features/quantitation/)quantifying 15 peaks in each spectrum arising from phosphocreatine (PCr), inorganic phosphates (Pi), ATP, and phosphomono- and diesters.16 The cerebral pH was determined from the frequency separation between Pi and PCr. Five consecutive spectra sets within the 31P time-series were averaged for 2 minutes and 30 seconds, which represent the metabolic baseline, first and second halves of breath-hold, as well as early and late metabolic recovery phases. This step was also to improve the signal-to-noise ratio. Thus, the time course of pH and of the ratios Pi/PCr, Pi/β-ATP, PCr/β-ATP, as well as the ratios of all 31P metabolites relative to (ATP + adenosine diphosphate), calculated from the mean of the γ- and α-ATP peaks, were obtained in the same 2 minutes and 30 seconds intervals as the ASL sequences. γ- and α-ATP peaks include the signals from nucleotide diphosphates and thus are expected as near-constant.
In a consecutive single-volume 1H-MR spectroscopy breath-hold experiment, ratios of the 1H-MR signals of N-acetylaspartate, total creatine, choline, and lactate were determined from an 8-mL volume in the left basal ganglia (point-resolved spectroscopy sequence localized spectra: TR 2 s; TE 0.14 s; 128 signal averages). Recording periods were split to match the ASL-MR imaging and 31P-MR spectroscopy phases of the breath-hold experiment.
Blood Gas Analysis
We drew 10 mL of venous blood before breath-hold, after 2 minutes and 30 seconds of breath-hold, at first breath after breath-hold, and after 2 minutes and 30 seconds of recovery. Immediate analysis (RAPIDLab 1265; Siemens, Erlangen, Germany) involved partial pressure of CO2 and oxygen (pO2), glucose, and lactate levels. With the exception of partial pressure of CO2, these parameters were shown to correlate well with arterial values.17
Statistics
Overall CBF variability was analyzed by a mixed linear model with the participant as a random factor, while differences between single time points were analyzed by a paired t test, which considers CBF differences compared with baseline (Δ CBF/CBF0) as the expression of CVR without division by a fixed rate of stimulus such as per unit CO2.18 The same tests were applied for the partial volume–corrected ASL-sCoV. MR spectroscopy parameters were analyzed in a mixed linear model and with the Pearson R correlation. The GM CBF in the vascular territories was analyzed separately by a paired t test to identify differences between the anterior and posterior circulation. The influence of age and experience with prolonged breath-holds estimated from cumulative breath-hold events of >2 minutes and 30 seconds was tested for correlation with CBF and ASL-sCoV using a mixed linear model with the participant as a random factor. Similarly, the relationships of the ASL parameters with SpO2 and HR were analyzed. Parametric testing occurred after testing for normal distribution. Generally, absolute CBF differences (Δ in mL/100 g/min) were used for descriptive statistics and statistical analyses, while ratios (in percentages) are mentioned for illustrative purposes.
Results
Cerebrovascular Reactivity
All participants showed a significant CVR by an increase in CBF until 5 minutes of breath-hold with a subsequent decline at recovery (P = .001 for all vessel territories, GM, and WM; On-line Table and Figs 1 and 2A, -B). While the CBF increase from baseline was substantial after 5 minutes (late breath-hold scan phase for the total GM ROI: mean Δ CBF, 18.3 ± 14.4 mL/100 g/min [+51.8%]; P = .001), it was only subtle within the first 2.5 minutes of breath-hold (total GM ROI: mean ΔCBF, 6.3 ± 11.1 mL/100 g/min [+17.8%]; P = .04). In 4 volunteers, early breath-hold-phase CBF was indeed lower than baseline CBF (mean, −22.4%; P = .01). Baseline-to-late recovery phase CBF differences were not significant (P = .55 for the total GM ROI). Return to baseline CBF or below occurred during the early recovery phase with a mild secondary CBF increase during the second 2 minutes and 30 seconds of recovery (P = .03 in total GM ROI).

Mean CBF during the breath-hold experiment. False color maps of cohort mean cerebral blood flow during 5-minute breath-hold. Selected transverse brain sections of the cohort mean CBF before, during, and after the 5-minute breath-hold challenge. The mean total GM CBF over all participants and over all phases was scaled to 60 mL/100 g/min. Likewise, the mean total WM CBF for all participants over all phases was scaled to 20 mL/100 g/min for WM.

Cerebral blood flow and partial volume–corrected relative spatial coefficient of variation dynamics. Baseline scan with normal ventilation starts at 0 minutes, the second and third measurement points represent 2 minutes and 30 seconds and 5 minutes of breath-hold, and the fourth and fifth scan phases at 7 minutes and 30 seconds and 10 minutes represent the recovery phase. A, CBF in gray matter voxels only (n = 15 participants, each represented by a color dot). B, The same for white matter only. C, Partial volume–corrected relative spatial coefficient of variation dynamics in gray matter. Y-axis: relative CBF spatial coefficient of variation defined as the ratio of the actual spatial coefficient of variation divided by the spatial coefficient of variation expected on the basis of anatomy (in arbitrary units [au]).
The absolute CBF and CVR of the anterior circulation (anterior cerebral artery, MCA) were, at all times, higher than in the posterior circulation (P = .001; On-line Table). The mean difference between CBF of the MCA and the posterior cerebral artery increased steadily during breath-hold from 8.8 ± 6.6 mL/100 g/min at baseline to 14.6 ± 6.1 mL/100 g/min at late breath-hold (P = .001).
During all scan phases, between-participant CBF variability showed a narrower range in the GM than in the WM as well as a lower overall CVR (CVR variability in GM: 83.8%–152.8% from baseline; CVR in WM: 74.2%–231.5% increase from baseline CBF; P = .001; On-line Table).
Spatial Coefficient of Variation
The ASL-sCoV varied over the course of the experiment (P = .001). In most cases (n = 12/15), there was an ASL-sCoV decrease between baseline and the early breath-hold phase (mean decrease: −30.0% ± 21.6%; P = .002; Fig 2C). The ASL-sCoV remained reduced during the 5-minute breath-hold (P = .81 for the difference between early and late breath-hold) and rose again during recovery (P = .01). There was no difference between the baseline and recovery phases (P = .29).
The range of GM ASL-sCoV among participants was smaller during breath-hold than during normal breathing: mean, 2.8% ± 1.0%; range, 1.9%–5.3% versus mean, 1.8% ± 0.4%; range, 1.5%–3.0%, for baseline versus early breath-hold phases, respectively. This range equates to a 35.7% decrease of ASL-sCoV variability among participants (P = .02).
Physiologic Correlations
The dynamics of SpO2 and HR are presented in the On-line Table. SpO2 correlated with CVR, with an estimated increase of CBF of 0.82 mL/100 g/min with each 1% SpO2 drop (95% CI, 0.6–1.1 mL/100 g/min; P = .001; Fig 3A, -B). HR was not correlated with CVR (P = .36; Fig 3C, -D). Similarly, age was not identified as an influential cofactor on CVR (P = .32).

Relationship of SpO2 and heart rate with CBF. A, Mean GM CBF (colored dots) and mean WM CBF (triangles) are both strongly correlated with absolute SpO2. Color encoding for A and C: green for baseline, orange for 2 minutes and 30 seconds breath-hold, red for 5-minute breath-hold, turquoise after 2 minutes and 30 seconds of recovery, and blue after 5 minutes of recovery. The same WM time points are right below. B, ΔCBF (here measured in GM) similarly correlates well to ΔSpO2 (all 75 single values). C and D, There is no strong correlation between heart rate and CBF in absolute values (C, mean) or ΔCBF and ΔSpO2 (D).
If one took the entire group of 15 freedivers into account, previous prolonged breath-hold events were not correlated with absolute CBF values (P = .56). However, there were 2 outlier participants (divers 11 and 13) with exceptionally extensive experience and comparatively high CBF. After their exclusion, a relationship between previous breath-hold experience and absolute CBF was found with 1000 previous prolonged breath-holds, reducing CBF in GM by 2.2 mL/100 g/min (95% CI, 0.7–3.7 mL/100 g/min.; P = .01; Fig 4) for the remaining 13 cases. This finding was similar for CBF in WM: One thousand previous breath-holds reduced WM CBF by 0.6 mL/100 g/min; 95% CI, 0.15–1.1 mL/100 g/min (P = .01). Mean CVR, however, as defined by the Δ CBF/CBF0, was not correlated with previous experience with prolonged breath-holds (P = .23). Similarly, ASL-sCoV was not correlated with SpO2, HR, age, or diver experience with prolonged breath-holds (P = .45, .53, .90, and .69, respectively).
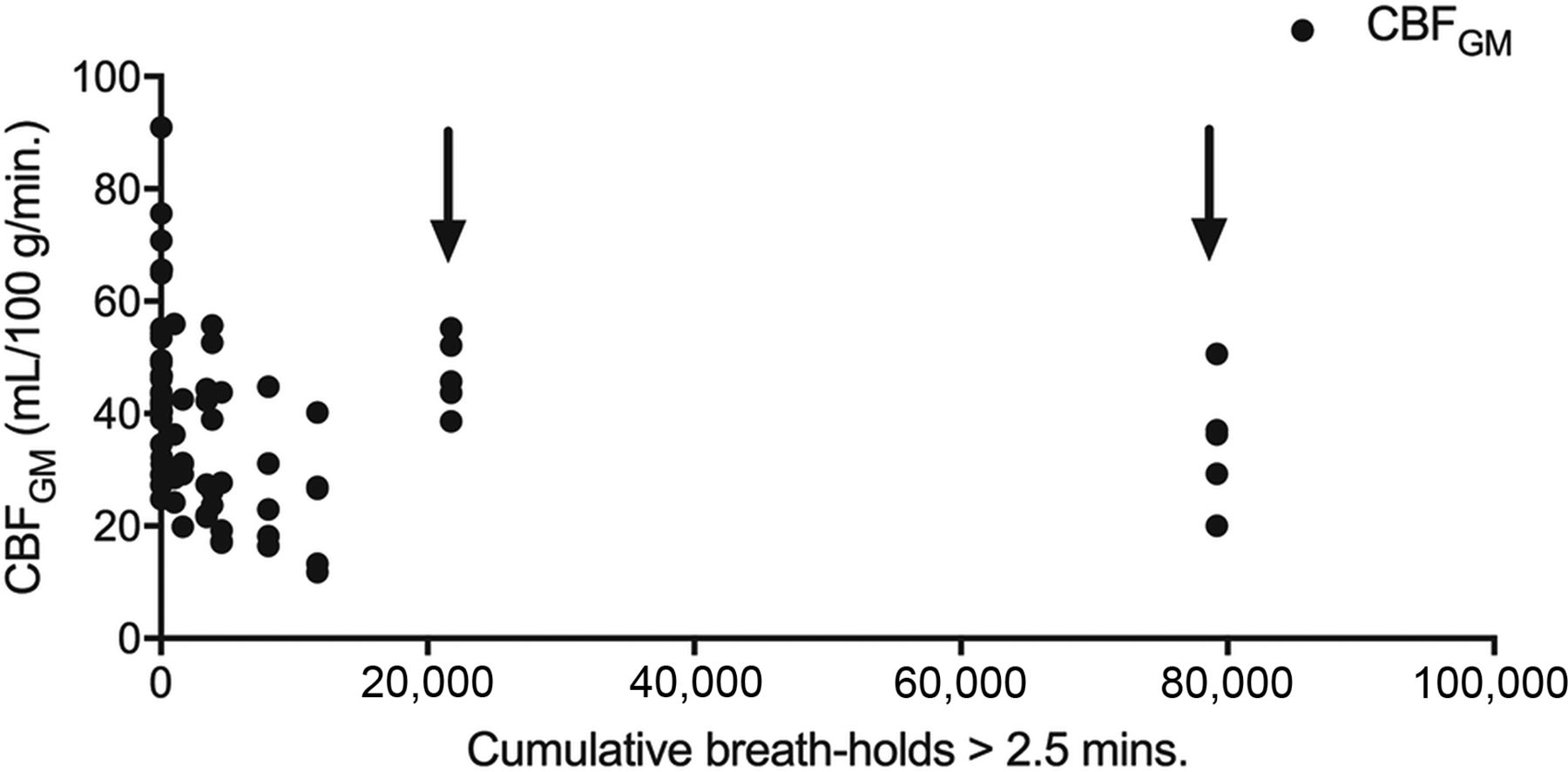
Scatterplot illustration of the relationship between cumulative breath-hold experience and CBF during the experiment. With the exception of 2 participants (divers 11 and 13, marked with arrows), there is a lower CBF during the experiment with more previous prolonged breath-hold events (cumulative breath-hold, >2 minutes and 30 seconds).
MR Spectroscopy and Blood Analyses
31P-MR spectroscopy (Fig 5A) revealed minor fluctuations in pH (Fig 5B) and ATP metabolites within physiologic ranges during the course of breath-hold (P = .07 for pH; P > .05 for dynamics in relative PCr; Pi; ATP-α, -β, -γ; and phosphomono- and diester levels). There was a small-but-significant decrease of the PCr / (ATP + adenosine diphosphate) ratio between baseline and late-phase breath-hold in the matched-pairs analysis of the individuals (−6.4%; P = .02; Fig 5C). PCr and β-ATP differences relative to their baseline values (ΔPCr and Δβ-ATP) were correlated with the differences in pH from baseline (R = 0.53; P < .001 for ΔPCr and R = 0.45; P = .003 for Δβ-ATP, respectively; Fig 5D).

Energy metabolites and blood gases in brain and venous blood. A, Sample 31P-MR spectrum of 1 participant averaged over 2 minutes and 30 seconds of the late breath-hold phase (above), displayed together with AMARES-fitted spectral components (below). B, Brain pH as assessed by 31P-MR spectroscopy is near-constant during the entire breath-hold experiment. C, The PCr / (ATP + adenosine diphosphate) ratio measured by 31P-MR spectroscopy slightly decreases during breath-hold. D, PCr and β-ATP differences (Δ) from individual baseline values correlate with the pH differences from baseline, indicating that small tendencies toward acidosis and ATP depletion occur during breath-hold. E, Venous blood analyses reveal significant hypoxemia and hypercapnia development (left axis in millimeters of mercury) during breath-hold with fast recovery. Blood glucose (in milligrams/deciliter, left axis) and lactate (in mmol/L, right axis) rose during breath-hold and did not return to baseline. All values are expressed as differences from baseline.
1H-MR spectroscopy never showed the CH3 doublet of lactate at 1.34 ppm in any of the participants. No significant changes in the levels of N-acetylaspartate, total creatine, or choline occurred (P > .05 for all).
Venous blood samples showed a development of hypoxemia and hypercapnia during breath-hold (On-line Table, Fig 5E). A significant partial pressure of CO2 increase was noted only at late breath-hold (P = .02), while pO2 had already dropped significantly after the early breath-hold phase (P = .002). Both parameters returned to baseline after breath-hold, while venous lactate and glucose levels increased until the end of the experiment during the recovery phase (P = .001 and .01, respectively; Fig 5E).
Discussion
This study provides 3 key findings. First, despite individual variability, freedivers show a relatively consistent and vessel-territory-specific CVR during a breath-hold challenge, which is measurable with ASL. Second, CBF and the ATT correlate ASL-sCoV deliver independent aspects of the cerebrovascular response to breath-hold. Finally, this study identified indicators for an influence of earlier experience with prolonged breath-hold on absolute CBF during the breath-hold challenge but not on CVR itself. Physiologic responses can apparently withstand the extreme biochemical challenge induced by a prolonged breath-hold of 5 minutes, which can be detected by ASL and MR spectroscopy.
Multiple methods exist to assess CBF, including ASL, phase-contrast MR imaging, PET, and Doppler sonography. Maximum breath-hold studies are rare, and CBF evaluations were, until now, exclusively performed with Doppler sonography, which revealed a continuous elevation of flow velocity in the MCA of around 100%.19,20 The mean maximum CBF increase after 5 minutes of breath-hold observed in this ASL study was 51.8%, which is very close to values observed in ASL studies using hypercarbic gas inhalation or short breath-hold, but indeed in some cases lower than values measured with ASL in a recent maximum breath-hold study (+107%).5,21,22 Previous studies comparing CBF assessments with different methods similarly revealed substantial intermethod differences in absolute CBF but otherwise confirmed a high correlation between methods. PET-estimated absolute CBF was, for example, consistently lower than phase-contrast MR imaging, while Doppler and ASL differed substantially in relative CBF change in a drug-stimulation trial.23,24 These differences are not surprising and can be explained by different influential factors acting on the respective flow parameters. While ASL can measure absolute CBF, Doppler provides flow velocity in a local vessel segment as a surrogate parameter for CBF. CO2 is a strong vasodilating agent in cerebral tissue causing an increase in CBF due to increased blood volume (CBV) based on the equation CBF = CBV / MTT. The blood flow velocity rises despite vessel dilation also due to reduced ATT. However, earlier Doppler studies revealed a strong neuromuscular response in proximal vessel segments of the anterior circulation, while more distal segments and the posterior circulation seemed less responsive.25,26 This may explain why Doppler-assessed breath-hold experiments identify a higher absolute and relative CVR, mostly in the M1 segments of the MCA, than most of those applying ASL, which captures perfusion in most distal and nonmuscular vascular segments.
A heterogeneous CVR between vessels was also observed in the present ASL study with lower CBF and CVR in the posterior cerebral artery territories. This finding can be explained by differences in labeling efficiency between vascular territories as well as the longer ATT in the posterior vascular territory. Microanatomic differences leading to a diverse autoregulation capacity between the anterior and posterior circulation as an underlying reason for a diverse CVR are, on the other hand, more controversially discussed even beyond the field of perfusion studies under extreme conditions.27,28 A recent Doppler-monitored breath-hold study supports our findings of a lower CVR in the posterior cerebral artery and suggests a different sympathetic activation between the 2 territories as an additional explanation beyond the labeling aspect, which needs to be considered in ASL.29 An awareness of a vessel-selective CVR is, however, crucial when interpreting ASL measurements in focal ischemic lesions after prolonged clinical conditions of apnea.
Another finding of this study is that CBF decreased during the first 2 minutes and 30 seconds of breath-hold in 4 of 15 participants. The counter-suggestive relatively higher CBF at baseline compared with later time points might be the effect of anticipation anxiety toward the upcoming breath-hold challenge.30 This well-known mental phenomenon among freedivers is currently not sufficiently addressed in sports physiologic research. Indeed, our own results may only suggest that some freedivers experienced an early sympathetic activation before the breath-hold challenge. Due to the limited temporal resolution of the ASL sequence, which delivers a mean CBF over each of the 2 minutes and 30 seconds phases, we cannot readily assess how long the reduction of CBF persists and when exactly an elevation of the CBF above took place during the early breath-hold phase in these 4 volunteers.
We further confirmed a difference between the cortical GM and the deep WM CVR, with the total GM CBF known to be at least 2 times higher than the WM CBF.31 Low WM signal is an obstacle for WM CBF assessment in ASL, despite the availability of background suppression.31 However, CVR differences between GM and WM tissue can be interesting because they may help to better understand morphologic findings in brain diseases such as obstructive sleep apnea syndrome.32 We noted a larger CVR variability and relative increase in WM than in GM, which could be explained by the later arrival of blood in the relatively more distal WM vessels. WM CBF is difficult to measure at baseline due to longer ATT. Direct ATT assessment was technically not possible as part of our experimental setting due to temporal restrictions and a resulting mono-post-labeling delay ASL sequence. However, our ATT approximation based on ASL-sCoV confirmed a breath-hold-induced decrease in ATT, which might increase the SNR of WM CBF, which inflates the measured ΔCBF to a certain extent.
For the interpretation of ASL-sCoV, it is viable to consider the methodologic peculiarities of ASL. An ATT increase will cause the ASL signal to appear in larger vessels, resulting in vascular artifacts. The ASL signal can, at the same time, decrease in areas with higher baseline ATT as in, for example, the perfusion watershed. These 2 effects both increase ASL-sCoV.8 However, ASL-sCoV has a theoretic lower limit attained when all labeled blood has arrived in the tissue and all vascular artifacts have already disappeared. Further ATT decrease beyond this limit will have only minimal effect on the ASL-sCoV. Reaching this lower limit of ASL-sCoV during the early breath-hold could explain the low ASL-sCoV variability in some participants. The ASL-sCoV decrease, however, appears to occur earlier than the CBF increase and levels off during the second breath-hold phase, while CBF further increases. An earlier response to breath-hold leading to decreasing ATT before the CBF increase in the late breath-hold phase can be the explanation, denoting that the effect is mainly vascular and perfusion changes as detected by ASL follow later. ASL-sCoV may be an earlier CVR marker of hypoxemia than absolute CBF.
CVR is predominantly triggered by changes in blood CO2, while, for example, hypoxemia detected by peripheral chemoreceptors is considered to play an independent-but-inferior role in cerebral vasodilation.33 While Willie et al20 already reported that O2 metabolism has a crucial influence on the breath-hold capability of freedivers, stressing the role of O2 for breath-hold tolerance and CVR, Cross et al19 could not confirm an influence of O2 on cerebral autoregulation in their prolonged breath-hold study. We identified falling O2 as an influential factor of CVR in this study, which can usually not be observed in CBF studies applying hypercarbic-normoxic gas despite otherwise comparable CVR between breath-hold and hypercarbic-normoxic gas studies.5,26 Due to the design of this study, which could not rely on arterial CO2 measurements, it remains, however, impossible to discern the relative contribution of hypercarbia and hypoxia to CBF increase. CVR increased faster in the second half of the breath-hold experiment in correlation with the secondarily faster SpO2 decline, which can be explained by pulmonary O2 stores that allow normal hemodynamic conditions during the first minutes of breath-hold. Cerebral near-infrared measurements in elite freedivers showed that cerebral O2 desaturation tends to occur within a mean of 175 ± 50 seconds, but not before, which supports our finding.34
In this study, CBF itself was not strongly correlated to HR. This finding does not allow concluding that CBF in breath-hold is not modulated by cardiac causes. It is, however, beyond the scope of this study to evaluate cardiac cofactors to CVR such as heart stroke volume or the diving reflex in detail. The factor age (an indirect measure of the cardiac and vascular influence on CVR) was assessed but was not associated with any of the flow parameters, which is not surprising considering that most volunteers in this study were younger than 40 years of age. Also, a selection bias of outstandingly healthy and well-trained freediver volunteers must be considered a further contributing factor.
A physiologic adaptation to breath-hold was another hypothesis to be tested in this study. A diminished CVR during hypercarbia/hypoxemia was reported for patients with chronic obstructive pulmonary disease as well as sleep apnea.35 In an attempt to investigate a similar association between experience with hypercarbic/hypoxemic states and CVR in freedivers, we estimated the total amount of previous prolonged breath-holds of the participants before participation in the current breath-hold experiment (defined as cumulative breath-holds of >2 minutes and 30 seconds in a lifetime). Our findings do not unconditionally corroborate that breath-hold experience accounts for an adaptation effect on CBF. First, CVR did not differ between more breath-hold-experienced freedivers and their less experienced counterparts. Second, by far, the 2 most prolonged breath-hold-experienced freedivers, who additionally stated a high frequency of longer breath-holds per training session, showed relatively high CBF values. However, for the remaining cohort, an association between experience and lower absolute CBF in all phases of the experiment could be observed, which may indicate that repetitive previous hypercarbia and hypoxemia have an acute cerebrovascular effect during a breath-hold challenge. Due to the limited cohort size, which also included freedivers normally specialized in shorter breath-holds, this interesting and also clinically relevant observation will need to undergo further critical evaluation in the future, favorably in a more homogeneous group regarding age and freediving specialization.
In prolonged breath-hold, the efficacy of the cerebrovascular but also the cardiac response to maintain a stable O2 supply to the brain despite decreasing availability is a crucial health aspect and can be assessed spectroscopically regarding energy metabolism. Direct noninvasive in vivo measurements of brain energy metabolism during prolonged breath-hold are extremely rare.22,36 Cerebral lactate accumulation or acidosis was observed in none of our participants, suggesting a sufficient compensation of limited O2 supply by recruitment of ATP stores and increasing CBF. However, we identified a substantial decrease in the PCr / (ATP + adenosine diphosphate) ratio during breath-hold. This can be interpreted as a compensatory PCr decrease to provide ATP by PCr hydrolysis as a consequence of declining O2 availability and reduced aerobic ATP production capacity in prolonged breath-hold.
Rising peripheral venous lactate levels during the breath-hold challenge in contrast to stable cerebral lactate stresses the shift toward a preferred cerebral O2 supply in breath-hold, including a CBF increase and, simultaneously, peripheral vasoconstriction. The rising glucose levels are likely a consequence of adrenaline-induced glucose mobilization and underline the exceptional metabolic and mental challenge of prolonged breath-hold.37 The correlative venous pO2 analyses documented pathologically low O2 levels after 5 minutes of breath-hold (down to 60% SpO2) and corroborated similarities of this breath-hold experiment with clinical settings of hypoxemia. On the other hand, untrained persons may encounter life-threatening consequences under these circumstances, while freedivers face hypoxemia under voluntary and trained conditions.
Maximum breath-hold without contact with water is a particular challenge for freedivers because regular training sessions mostly include water immersion, which causes an augmented diving reflex. Only very few participants fulfilled the inclusion criteria and were able to perform sufficient breath-hold in the noisy MR imaging environment. Multiple candidates declined due to the lack of silence, or they would not volunteer for additional MR spectroscopy breath-hold experiments, thus the limited cohort size. Furthermore, participants did not tolerate a CO2 mask, which interfered with their meditative state. We therefore decided to analyze venous CO2. However, while venous CO2 can be used to confirm hypercapnia, it is, however, too variable to study the CO2 influence on CVR due to known substantial deviations from arterial CO2 in the brain.38 For this reason, we can also only assume that all normoventilating volunteers started at normal CO2 blood levels in the experiment. The unavailability of these otherwise valuable data limits the evaluation of the physiologic processes behind our observations.
Conclusions
This study revealed that experienced freedivers develop a CVR, which is sufficient to maintain a physiologic cerebral energy metabolism even during a prolonged breath-hold period of 5 minutes and severely diminishing blood O2. Furthermore, ASL parameters, which are determined by blood flow and vessel diameter alterations alike, serve as excellent candidate MR imaging parameters to reveal this response, while 31P-MR spectroscopy revealed its utility to dynamically study acute changes in cerebral energy metabolism. ASL may also provide evidence for long-term adaptation of cerebral vasculature following repetitive hypoxia-hypercapnia. Imaging and metabolic findings of the present freediver study can be used to better understand CVR during hypoxia-hypercapnia in critical care and sleep apnea conditions.
Acknowledgments
The authors acknowledge Dipl.-Phys. Jürgen Gieseke for his assistance with the ASL sequence implementation, and we thank all freedivers for their outstanding effort to make this study possible.
Footnotes
Vera C. Keil, Lars Eichorn, and Henri J.M.M. Mutsaerts contributed equally to this work.
Disclosures: Burkhard Mädler—UNRELATED: Employment: Philips Healthcare, Comments: regular salary; Kim van de Ven—UNRELATED: Employment: Philips Healthcare. Comments: regular salary.
Paper previously presented, in part, at: 99th German Röntgen Congress, May 27, 2017 (No. 4735), Leipzig, Germany; and Joint Annual Meeting of the International Society for Magnetic Resonance in Medicine and the European Society for Magnetic Resonance in Medicine and Biology, June 16–21, 2018; Paris, France.
References
- Received December 29, 2017.
- Accepted after revision July 19, 2018.
- © 2018 by American Journal of Neuroradiology





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}